| Journal of Food Bioactives, ISSN 2637-8752 print, 2637-8779 online |
| Journal website www.isnff-jfb.com |
Original Research
Volume 17, March 2022, pages 34-48
In silico investigation of molecular targets, pharmacokinetics, and biological activities of chicken egg ovalbumin protein hydrolysates
Toluwase Hezekiah Fatokia, *, Rotimi E. Alukob, c, Chibuike C. Udenigwed, e
aDepartment of Biochemistry, Federal University Oye-Ekiti, PMB 373, Oye-Ekiti, Ekiti State, Nigeria
bDepartment of Food and Human Nutritional Sciences, University of Manitoba, Winnipeg, MB R3T 2N2, Canada
cRichardson Centre for Functional Foods and Nutraceuticals, University of Manitoba, Winnipeg, MB R3T 2N2, Canada
dSchool of Nutrition Sciences, Faculty of Health Sciences, University of Ottawa, Ottawa, ON K1H 8M5, Canada
eDepartment of Chemistry and Biomolecular Sciences, Faculty of Science, University of Ottawa, Ottawa, ON K1N 6N5, Canada
*Corresponding author: Toluwase Hezekiah Fatoki, Department of Biochemistry, Federal University Oye-Ekiti, PMB 373, Oye-Ekiti, Ekiti State, Nigeria. E-mail: toluwase.fatoki@fuoye.edu.ng
DOI: 10.31665/JFB.2022.17302
Received: March 6, 2022
Revised received & accepted: March 26, 2022
| Abstract | ▴Top |
Food-derived bioactive peptides are promising ingredients for developing functional foods and nutraceuticals due to their putative safety, low cost, and multiple health benefits. Chicken egg is considered a major source of dietary protein, lipids, vitamins, and minerals but is also highly allergenic. The aim of this work was to investigate the inherent bioactive properties of chicken ovalbumin peptides using in silico approaches. Ovalbumin was in silico hydrolyzed with gastrointestinal proteases (chymotrypsin, trypsin, and pepsin) and results indicated cleavage of the most allergenic protein with an overall 36.62% theoretical degree of hydrolysis, consisting of 132 fragments of which 65 were di-, tri-, tetra- or oligopeptides. The most represented biological targets obtained for these peptides include HLA class I histocompatibility antigen A-3, E3 ubiquitin-protein ligase XIAP, and angiotensin-converting enzyme. Notably, peptides AIVF and AVL were found to have multi-target potentials. Gene enrichment analysis showed interaction of these peptides with some kinases and transcription factors. Overall, results from binding affinity, pharmacokinetics and physicochemical properties, and therapeutic activity showed that PGF, SSL, GGL, AVL, VY, and IL are promising peptide candidates for further studies. These results are important in the design of peptide-based functional foods and therapeutic products devoid of allergenic property of ovalbumin.
Keywords: Ovalbumin; Hydrolysate; Bioactive peptides; Docking; Pharmacokinetics; Therapeutic potential; In silico
| 1. Introduction | ▴Top |
Proteins are integral food components that provide nutritional, technological, and functional properties. Bioactive peptides (BAPs) are amino acid chain fragments that are generated during proteolytic cleavage or maturation of functional proteins and exhibit similar or unrelated biological properties in comparison to the parent protein (Iavarone et al., 2018). Food-derived BAPs are short sequences of 2–20 amino acids that are embedded within the primary structure of dietary proteins (Manzanares et al., 2019). Normally, these peptide sequences are inactive within the precursor protein but can be activated during food processing or enzymatic hydrolysis (Daliri et al., 2017). The precise functions of these peptides depend on their amino acid composition, sequence, and chain length in addition to physicochemical properties (Sánchez and Vázquez, 2017). Food-derived BAPs are promising ingredients for developing functional foods and nutraceuticals due to their putative safety, low cost, and multiple health benefits (Udenigwe and Aluko, 2012). A growing body of scientific evidence indicates that peptides regulate several physiological processes and act as antidiabetic, antihypertensive, antimicrobial, antioxidant, anticancer, and immunomodulatory agents (Antony and Vijayan, 2021; Chalamaiah et al., 2018). Consequently, the study of food protein hydrolysates has become a popular area in biochemical and nutritional research (Famuwagun et al., 2020, 2021; Wu et al., 2020; Girgih et al., 2015).
BAPs have been isolated from food sources, including animal (milk, egg, cheese, yoghurt, and meat), vegetable (pulses, lentils, nuts, wheat, rice, oat, and flaxseeds), and marine organisms (fish, tuna, salmon, squid, oyster, and crabs) proteins (Antony and Vijayan, 2021). BAPs are released from food proteins by exogenous or endogenous proteases, or microbial fermentation (Xu et al., 2019). In vitro, in vivo or in silico enzymatic hydrolysis can also be used for BAPs preparation. To produce BAPs with specific activity, proteases with broad cleavage specificity are used for proteolysis, e.g., bromelain, ficin, and papain from plant sources; chymotrypsin, pepsin, trypsin, and pancreatin from animal sources; and Alcalase, collagenase, Flavourzyme, Neutrase, pronase, proteinase K, and subtilisin A from microbial sources (Jakubczyk et al., 2020; Yada, 2017).
Eggs are widely consumed globally and Gallus gallus (chicken) egg is one of the most preserved natural products that is considered a major source of proteins, lipids, vitamins, and minerals (Abeyrathne et al., 2013). The egg yolk consists of lipovitellins (36%), livetins (38%), phosvitin (8%), low-density lipoproteins (17%), and carotenoids (1%), which impart the yellow color (Nolasco et al., 2019; Abeyrathne et al., 2013). The constituent major proteins in egg white are ovalbumin (60–65%), ovotransferrin (12%), ovomucoid (11%), lysozyme (3.5%), and ovomucin (3.5%), while avidin (0.05%), cystatin (0.05%), ovomacroglobulin (0.5%), ovoflavoprotein (0.8%), ovoglycoprotein (1.0%), and ovoinhibitor (1.5%) are the minor proteins (Guerin-Dubiard et al., 2006; Kovacs-Nolan et al., 2005; Stein et al., 1991).
Egg white ovalbumin belongs to the serpins superfamily found in animals, plants and viruses, and they probably evolved from a serine protease inhibitor; example of serpins include plasma inhibitors, hormone-binding globulins, and angiotensinogen (Stein et al., 1991). Ovalbumin is a glycoprotein with a molecular weight of ∼45 kDa. It lacks the recognized protease inhibitory activity despite having 30% sequence identity with antitrypsin and similarity with other functional inhibitors of the serpin family (Stein et al., 1991). Chicken ovalbumin (Gene ID: SERPINB14) consists of 386 residues and functions as a storage protein and mediator of biological processes such as amino acid and ion homeostasis, and negative regulation of endopeptidase activity (https://www.uniprot.org/uniprot/P01012).
Several tools are now available for efficient proteomic studies (Guerin-Dubiard et al., 2006), especially, bioinformatics methods for predicting the potential sequence of bioactive peptides. In silico methods are used to study bioactive peptides from various plants and animals due to their cost-effectiveness, timeliness, and dependability (Agyei et al., 2018). For instance, a whole peptidomics and bioinformatics study of chicken egg white revealed the presence of 43 novel antidiabetic peptides based on homologous sequence motifs identified in other food-derived peptides (Arena et al., 2020). According to Majumder and Wu (2010), novel peptides could be identified by the use of quantitative structure-activity relationship (QSAR) and bioinformatics. An overview of the application of in silico methods for the generation of food protein-derived BAPs has been provided in recent reviews by Agyei et al. (2018) and FitzGerald et al. (2020). The current research work fills the gap in knowledge in the previous studies on chicken egg white (Zhao et al., 2020; Salim and Gan, 2020) by addressing the pharmacokinetic profiles, molecular targets affinity, and biological activities of potential BAPs. Therefore, the aim of this work was to investigate the inherent pharmacological properties of ovalbumin using in silico approaches to release and identify BAPs that could be used to formulate functional foods and therapeutic products.
| 2. Materials and methods | ▴Top |
2.1. Protein hydrolysis and bioactivity assessment
The sequence of ovalbumin from Gallus gallus (chicken), with the gene name: SERPINB14 and UniProt ID: P01012, was obtained from the protein sequence database UniProt (www.uniprot.org) in FASTA format. Enzymatic hydrolysis of ovalbumin was achieved using BIOPEP-UWM (http://www.uwm.edu.pl/biochemia/index.php/en/biopep), which performs enzymatic hydrolysis and bioactivity prediction based on known BAPs in the database (Minkiewicz et al., 2019). Simulated combined digestion was done in BIOPEP-UWM using gastrointestinal (GI) enzymes, such as chymotrypsin (EC 3.4.21.1), trypsin (EC 3.4.21.4), and pepsin, pH 1.3 (EC 3.4.23.1). The theoretical degree of hydrolysis (DH) was calculated from the equation: TDH = [d/D] × 100%, where d is number of hydrolyzed peptide bonds and D is total number of peptide bonds in a protein chain. Potential biological activity of the released peptides was determined using the appropriate BIOPEP-UWM tool.
2.2. Target prediction
Target prediction was conducted using the sequence of 65 peptides obtained from the in silico enzymatic hydrolysis (Table s1) as well as the sequence of 6 selected peptides with potential biological activity from the result of initial screening on BIOPEP-UWM to serve as control. The amino acid sequence was converted to a structure format called Simplified Molecular Input Line Entry Specification (SMILES) using PepSMI (https://www.novoprolabs.com/tools/convert-peptide-to-smiles-string) as described by Fatoki et al. (2020a). In silico prediction of target was done using SwissTargetPrediction (STP) (http://www.swisstargetprediction.ch/), where Homo sapiens was selected as the target organism (Diana et al., 2019), and selection was made based on a minimum of 25% probability. Lastly, Super-PRED (SP) was used to predict targets that are not analyzable on STP (https://prediction.charite.de/subpages/target_prediction.php).
2.3. Peptide-peptide docking simulations
Based on the results obtained from the target prediction, protein-peptide docking was done using HPEPDOCK (https://huanglab.phys.hust.edu.cn/hpepdock/) (Zhou et al., 2018) at default setting, using the protein structure in PDB format and the peptide sequence in FASTA format. PyMOL software v2.0.7 was used for visualizing the three-dimensional interactions of the protein-peptide complexes. Binding energy of the protein-peptide complex was calculated using Prodigy (https://bianca.science.uu.nl/prodigy/) (Xue et al., 2016).
2.4. Target gene expression analyses
Twenty genes were obtained from target prediction results for 25 ovalbumin peptides. The gene IDs (FPR2, CTSB, CAPN1, FURIN, CFB, HLA-A, FNTA, FNTB, PTGS2, ACE, OPRM1, ITGAV, SLC5A1, HMGCR, BACE1, YARS1, DPP4, XIAP, TACR1, and OPRD1) were the basis for expression network analyses (transcription factor enrichment analysis, protein-protein interaction network expansion, and kinase enrichment analysis), using eXpression2Kinases (https://maayanlab.cloud/X2K/) (Clarke et al., 2018), where ‘human’ was listed as the background organism.
2.5. Pharmacokinetics and physicochemical properties prediction
The active peptides (peptides with predicted target) were subjected to in silico Absorption, Distribution, Metabolism, and Excretion (ADME) screening on SwissADME (http://www.swissadme.ch/) at default settings (Diana et al., 2017). Potential toxicity of the peptides was predicted by virtual scanning using ToxinPred (https://webs.iiitd.edu.in/raghava/toxinpred/multi_submit.php; Gupta et al., 2013). Hemolytic or red blood cell (RBC) lysing potential of the peptides was estimated using HemoPI (https://webs.iiitd.edu.in/raghava/hemopi/), which was also used to calculate the theoretical isoelectric point (pI), hydrophobicity, hydropathicity, hydrophilicity, and net charge at pH 7.
2.6. Prediction of therapeutic potential of the peptides
Therapeutic potential, including anticancer, antimicrobial, antiviral, antioxidant, anti-inflammatory. antihypertensive and anti-diabetic activities, of the ovalbumin peptides was conducted using (i) iAMPpred (http://cabgrid.res.in:8080/amppred/; Meher et al., 2017) for antimicrobial (antibacterial, antiviral and antifungal peptides), (ii) PreAIP (http://kurata14.bio.kyutech.ac.jp/PreAIP/; Khatun et al., 2019) for anti-inflammatory properties, (iii) AntiCP server virtual screening (https://webs.iiitd.edu.in/raghava/anticp/multi_pep.php) for anticancer properties (Tyagi et al., 2013), and (iv) AodPred web-server (http://lin-group.cn/server/AntioxiPred, Feng et al., 2016) for antioxidant peptides. Peptide sequence in FASTA format was used for all the analyses.
| 3. Results | ▴Top |
3.1. Protein hydrolysis
In silico hydrolysis of ovalbumin yielded 132 fragments from the combined actions of GI proteases (chymotrypsin, trypsin, and pepsin), which hydrolyzed 141 of the 385 peptide bonds in the protein (Figure 1) resulting in a theoretical degree of hydrolysis of 36.62%. Several biologically active peptides were screened on BIOPEP-UWM for intact (unhydrolyzed) and hydrolyzed ovalbumin sequences having potential antioxidative, antihypertensive, antidiabetic, hypolipidemic and kinase activities, as shown in Tables 1 and 2. Only 65 of the 132 fragments from hydrolyzed ovalbumin with two or more amino acid residues and six most active peptides from the unhydrolyzed screening were selected for further analyses (Table s1).
 Click for large image | Figure 1. Fragments of ovalbumin released by the action of the GI enzymes, chymotrypsin, trypsin, and pepsin. |
 Click to view | Table 1. Predicted peptides (with minimum of three amino acid residues) of intact ovalbumin (UniProt ID: P01012) with potential biological activity from BIOPEP-UWM |
Click to view | Table 2. Ovalbumin hydrolysate peptides that were predicted to be biologically active on BIOPEP-UWM |
3.2. Target prediction
Eighteen targets were selected for further studies based on a minimum score of 25% probability as shown in Table 3. After analysis, the most represented targets obtained (with their UniProt ID) were HLA class I histocompatibility antigen A-3 (P04439), E3 ubiquitin-protein ligase XIAP (P98170), and angiotensin-converting enzyme (P12821), while peptides that have multi-target potentials were AIVF and AVL. Peptide CPIAIM had the highest (60%) probability of target for protein farnesyltransferase (P49354, P49356); LPF (90%), AIVF (85%) and PIL (75%) for E3 ubiquitin-protein ligase XIAP (P98170); GGL (70%) for cyclooxygenase-2 (P35354); PIL (75%) for dipeptidyl peptidase IV (P27487); and VY (60%) for tyrosyl-tRNA synthetase (P54577). The hydrolysate peptides GDSIEAQCGTSVN, QPSSVDSQTAM, and EVVGSAEAGVDAASVSEEF could not be analyzed on SwissTargetPrediction due to their bulkiness, thus Super-PRED (SP) was used. The result showed that the three bulky peptides could target cathepsin D (P07339), delta opioid receptor (P41143), caspase-1 (P29466), cyclooxygenase-2 (P35354), and others.
Click to view | Table 3. Protein targets of ovalbumin protein hydrolysates predicted from SwissTargetPrediction |
3.3. Target gene expression network
Results from the enrichment analysis based on the hypergeometric (−log10) p-value (Figure 2) showed that the peptides targeted the expression of kinases (Gene ID, UniProt ID), including casein kinase II subunit alpha (CK2ALPHA/CSNK2A1, P68400), mitogen-activated protein kinases (MAPK1/ERK2, MAPK3/ERK1, and MAPK14; P28482, Q16539), and homeodomain-interacting protein kinase 2 (HIPK2, Q9H2X6). In addition, the peptides targeted the expression of transcription factors, including signal transducer and activator of transcription 3 (STAT3, P40763), nuclear factor erythroid 2-related factor 2 (NFE2L2, Q16236), protein C-ets-1 (ETS1, P14921), erythroid transcription factor (GATA1, P15976), transcription factor PU.1 (SPI1, P17947), interferon regulatory factor 8 (IRF8, Q02556), transcriptional regulator ERG (ERG, P11308), and polycomb protein SUZ12 (SUZ12, Q15022). The lower the hypergeometric p-value, the better the enrichment.
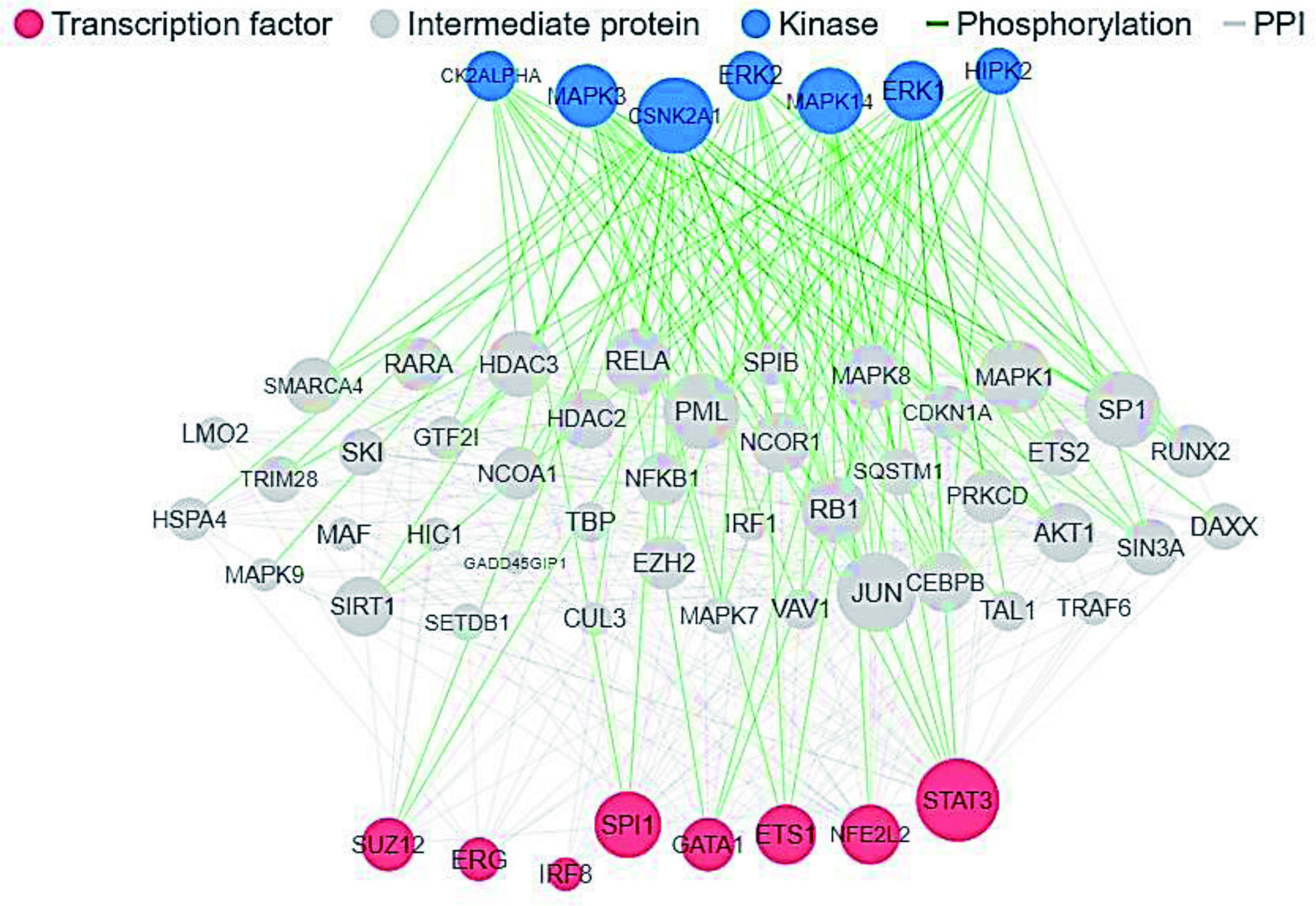 Click for large image | Figure 2. eXpression2Kinases Network showing overall interactions of intermediate proteins, kinases and transcription factors with high hypergeometric (−log10) p-value. |
3.4. Peptide-peptide docking simulations
The best models obtained from HPEPDOCK scores (Table s2) were used for calculation of binding free energy (ΔG) and dissociation constant on PRODIGY. Figure 3 shows the binding of selected peptides (number) to protein targets (alphabet), visualized using PyMOL. The binding free energy result of the protein-protein docking for each of the ovalbumin peptides and their corresponding targets is shown in Table 4. The results of this study showed that peptide SGISSAESL has the highest binding free energy (ΔG, −9.6 kcal.mol−1) for HLA class I histocompatibility antigen A-3 (PDB ID: 1AKJ), followed by GSIGAASM (ΔG, −8.5 kcal.mol−1). GIIR, PGF and AVL have highest binding free energy (ΔG, −8.9 kcal.mol−1) for furin (PDB ID: 6HLD), angiotensin-converting enzyme (PDB ID: 4C2N) and dipeptidyl peptidase IV (PDB ID: 3Q8W) respectively. QTAADQAR has a binding free energy (ΔG) of −8.9 kcal.mol−1 for integrin alpha-V/beta-3 and alpha-5/beta-1 (PDB ID: 4O02) while GITDVF has a binding free energy (ΔG) of −8.4 kcal.mol−1 for beta-secretase 1 (PDB ID: 2OHM).
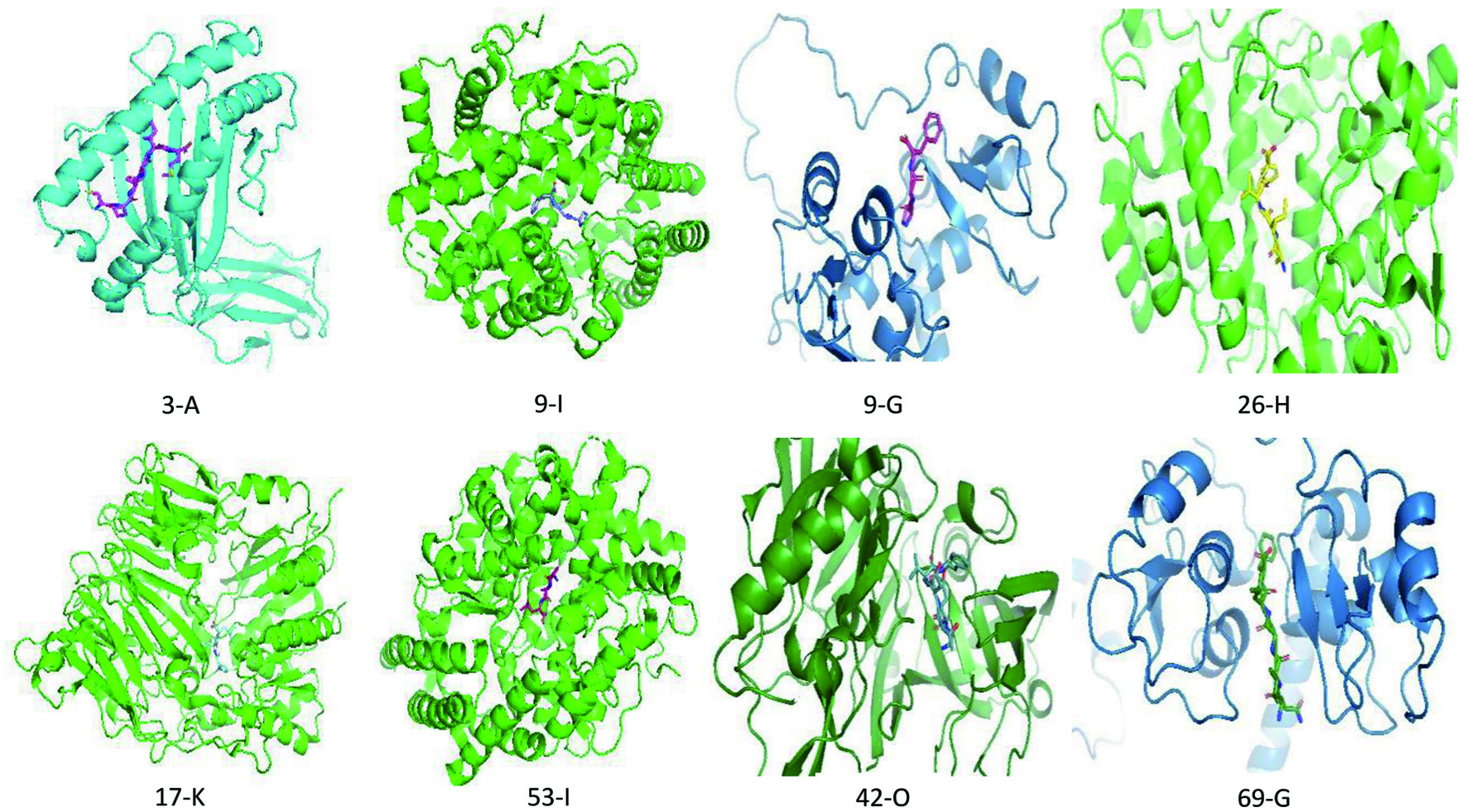 Click for large image | Figure 3. Binding interaction between selected peptides (number) and protein targets (alphabet) as shown in Table 4 and visualized using PyMOL. |
Click to view | Table 4. Predicted binding energy and dissociation constant of the protein-protein complexes obtained from PRODIGY |
3.5. Pharmacokinetics and physicochemical properties
The results in Table 5 show that peptides PGF, PIL, GGL, AVL, VY, IL and LPF have high gastrointestinal absorption (GA) rate. PGF, SSL, GGL, AVL, VY, IL and some other peptides were predicted not to be substrates for P-glycoprotein (P-gp). None of the peptides in this study was predicted to cross the blood-brain barrier or inhibit cytochrome P450 enzymes. The bioavailability score (BS) of PGF, SSL, PIL, GGL, AIVF, AVL, VY and some others were 0.55. QTAADQAR and SGISSAESL have low BS but high synthetic accessibility (SA). The results in Table 6 show that all the peptides in this study are non-toxic and non-hemolytic, and that PGF, SSL, PIL, GGL, QTAADQAR, AIVF, AVL, VY, IL and some other peptides have zero net charge.
Click to view | Table 5. Predicted pharmacokinetics properties of ovalbumin protein hydrolysates |
Click to view | Table 6. Predicted physicochemical properties of ovalbumin protein hydrolysates |
3.6. Therapeutic potential of the peptides
Table 7 shows that peptides CPIAIM, PGF, PIL, GGL, GIIR, QIGL, IL and LPF have antimicrobial properties, including anti-bacterial, antiviral, and antifungal activities. In addition, GSIGAASM, GAK, GGL, GIIR and GITDVF were predicted to have no anticancer activity. Only GIIR and QIGLF were predicted to have anti-inflammatory activity, while all of the peptides were predicted to have no antioxidant activity.
Click to view | Table 7. Predicted therapeutic potential of the peptides |
| 4. Discussion | ▴Top |
In silico hydrolysis of ovalbumin by the three GI proteases yielded hydrolysates containing several di-, tri-, tetra-, and oligo peptides (Figure 1). The screening results show the release of several peptides with potential therapeutic activities. The potential allergen peptide sequence (f323-339) was hydrolyzed by the GI protease treatment of ovalbumin in this study, and this would prevent production of interleukin-4 (IL-4) and the subsequent T-helper 2 (Th2) cytokine responses, which play critical roles in inducing food allergy inflammation (Nakajima-Adachi et al., 2006). Immunotherapy with hydrolysates of ovalbumin (OVA) and egg white (EW) produced with pepsin was reported to stimulate tolerance development in BALB/c mice allergic to EW more efficiently than treatment with the intact allergens, based on the induction of Treg (T) cells (Lozano-Ojalvo et al., 2017, 2019).
The T cell receptor (TCR) cross-reactivity between major histocompatibility complex II (MHCII)-binding to self and foreign peptides, such as nonamer peptides within f327-338 of ovalbumin, could influence the naïve CD4+T cell repertoire and autoimmunity (Nelson et al., 2015). Allergen peptide sequence f323-339 has been found to be a dominant determinant of CD4+T cells from egg allergy patients (Shimojo et al., 1994). CD4+T cells critically regulate the antigen-specific immune responses in food allergy or tolerance induction by recognizing the epitope peptides presented by MHC class II molecules (Nakajima-Adachi et al., 2012; Blazquez et al., 2010). Five distinct regions of chicken egg ovalbumin that contain dominant allergic immunoglobulin E (IgE) epitopes include L38-T49, D95-A102, E191-V200, V243-E248 and G251-N260 (Mine and Rupa, 2003). The hydrolysis of these regions by GI enzymes in this study produced several new active and inactive peptides (Figures s1 and s2). It is thought that only hydrolysis can completely eliminate the allergenic potential of a protein (Verhoeckx et al., 2015; 2019). Therefore, allergic reaction to ovalbumin in humans could be a result of defect in the structure and function of GI proteases. This is in line with a previous hypothesis linking resistance to digestion and allergenicity, in terms of defect in protease secretion and changes in the pH of gastric acid in disease state (Untersmayr and Jensen-Jarolim, 2008; Bernasconi et al., 2006; Untersmayr et al., 2003; Astwood et al., 1996). A previous study on intact ovalbumin immunotherapy subcutaneously performed in ovalbumin (OVA)-sensitized mice model of allergic asthma showed significantly reduced OVA-specific IL-4 and IL-5 production, and increased serum levels of OVA-specific IgG1, while OVA-specific IgG2a and IgE levels were not affected (Janssen et al., 1999).
The results of this study (Table 3) showed that most of the ovalbumin hydrolysate dipeptides obtained were not predicted to be bioactive against targets at a minimum of 25% probability. The prediction provided 18 protein targets in humans that may be modulated by the selected hydrolysate peptides of ovalbumin. These protein targets are involved in several disease processes, including hypertension (angiotensin-converting enzyme); immune diseases (beta-secretase 1, cathepsin B, calpain-1, complement factor B, and HLA class I histocompatibility antigen); diabetes (dipeptidyl peptidase 4, and sodium/glucose cotransporter 1); microbial infection (furin, integrin alpha-V/beta-6); and inflammation (prostaglandin G/H synthase 2 or cyclooxygenase 2, HMG-CoA reductase, protein farnesyltransferase, E3 ubiquitin-protein ligase XIAP, and tyrosine-tRNA ligase). In this study, peptides GSIGAASM, CPIAIM, SAL, GAK, SSL, SGISSAESL, AVL, which targeted HLA class I histocompatibility antigen A-3, all have hydrophobic amino acids at the position before the anchor residues (Met, Leu or Lys) at the C-terminus. This study showed that peptides VVR and GIIR had the same Arg at the C-terminus, and they can bind to furin. Peptides PGF, AIVF, LPF and QIGLF, contained the amino acid Phe at the C-terminus, and they can bind to angiotensin converting enzyme (ACE). Peptides PIL, QIGL, AVL, possessed the amino acid Leu at the C-terminus, and they can bind to DPP-IV to possibly exert mild inhibitory effects based on their calculated dissociation constant (kd).
Ovalbumin is a well-known source of peptides that possess antihypertensive, anticancer, anti-inflammatory and antioxidant activities (Nolasco et al., 2019) as shown in Tables 1 and 2. Previous works have reported that hydrolysis of egg white ovalbumin with pepsin produced peptides, of which two peptides, YREERYPILRADHPFL and IVF, showed strong ACE-inhibitory activities, vasodilation effect, and reduction of blood pressure in hypertensive rats (Miguel et al., 2007; Miguel and Aleixandre, 2006). Furthermore, ovalbumin hydrolyzed with pepsin, trypsin, and α-chymotrypsin produced seven angiotensin converting enzyme (ACE)-inhibiting peptides LKA, LKP, LAP, IKW, FQKPKR, FKGRYYP, and IVGRPRHQG (Fujita et al., 2000). Some of the peptides produced from ovalbumin not only showed strong ACE-inhibitory effects but also lowered blood lipid levels (Manso et al., 2008). Three other ovalbumin peptide sequences, RVPSL, TNGIIR and QIGLF, exhibited high ACE-inhibitory activity with inhibitory concentration that reduced enzyme activity by 50% (IC50) values of 20, 70, and 75 μM, respectively (Liu et al., 2010; Yu et al., 2011a, 2012a). However, some of the peptides shown in Table 1, specifically EVSGL, QITKPN, LEPINF, ANENIF, AEAGVD, EAGVD, DHPFLF, HAEIN and QIGLF exhibited α-amylase and α-glucosidase inhibitory activities with high (>150 μM) IC50 values, and thus did not possess anti-diabetic activity (Yu et al., 2011b; 2012b).
The network analysis of target genes (Figure 2) shows overall effect of ovalbumin hydrolysate peptides obtained in this study on multiple kinases and transcription factors when ingested by humans. Polycomb protein SUZ12 (UniProt ID: Q15022) functions in biological processes that involve response to retinoic acid, spinal cord development, negative regulation of tyrosine phosphorylation of STAT protein, and negative regulation of chemokine production (Cunningham and Duester, 2015; Corley and Kroll, 2015). Martínez-Blanco et al. (2020) reported that ovalbumin peptides obtained with pepsin enhanced the retinoic acid pathway on dendritic cells by activation of aldehyde dehydrogenase and transforming growth factor beta (TGF-β), through Toll-like receptor (TLR) interactions. This confers upon dendritic cells the ability to upregulate interleukin 10 (IL-10) as well as other tolerance-promoting mediators downstream of TRL signaling, such as IL-27, IL-33, Notch ligands, OX40L, and transcription factors such as Interferon regulatory factor 4 and 8 (IRF4 and IRF8). IRF8 (Uniprot ID: Q02556) is a transcription factor that binds to the upstream regulatory region of type I interferon (IFN) and IFN-inducible MHC class I genes. It mediates several biological functions such as defense response to bacterium and protozoan, positive regulation of interleukin-12 production, and transcription by RNA polymerase II (Berghout et al., 2013).
Results of this study also revealed homeodomain-interacting protein kinase 2 (HIPK2) as one of the key kinases modulated by ovalbumin hydrolysate peptides (Figure 2). HIPK2 (UniProt ID: Q9H2X6) is a serine/threonine-protein kinase involved in transcription regulation, p53/TP53-mediated cellular apoptosis, and cell cycle regulation. It is involved in the regulation of eye size, lens formation and retinal lamination during late embryogenesis (Inoue et al., 2010). Signal transducer and activator of transcription 3 (STAT3) acts in biological processes that involve glucose, temperature, and energy homeostasis, and inflammatory response, to mediate cellular responses to interleukins, leptins, peptide hormones, and other growth factors. Studies have shown that a specific STAT3 peptide aptamer decreased the expression level of Bcl-xL protein within the entire STAT3 pathway, as well as inhibition of STAT3 signaling, which led to the apoptosis in tumor cells (Borghouts et al., 2008; Nagel-Wolfrum et al., 2004).
Protein–peptide docking predicts the complex structure by sampling possible peptide binding conformations and ranking the putative complexes with an energy scoring function (London et al., 2013). The key concept of molecular docking is to develop an appropriate solution to elucidate the minimum free energy (ΔG) of interaction per mole of ligand (Pagadala et al., 2017). The molecular docking results showed the binding affinity of the ovalbumin peptides to their corresponding target proteins. Binding free energy (ΔG) scores of less than −5.0 kcal.mol−1 show good binding affinity (Figure 3). The smaller the dissociation constant, the better the affinity between the ligand and the protein. A previous study has shown that PGF and CPIAIM interacted with ACE at H383 and E411 from the active site residues, and also at Q281, H353, H383, E411, F457, K511, H513, Y520 and Y523 from the stabilizing residues, while PIL and GGL interacted only with ACE stabilizing residues (Salim and Gan, 2020). Similarly, peptides ADF, CDR, MIR, FGR, and FK have been found to interact with the ACE active site residues Lys511 and His353 (Zhao et al., 2020).
The GI epithelium interacts with food proteins after modification with gastric acid, gastric and pancreatic enzymes, and brush-border proteases, leading mainly to the production of peptides and amino acids, which are absorbed depending on size, polarity and shape (Snoeck et al., 2005). Overall, the results of ADMET in this study (Table 5) showed that most of the peptides have low gastrointestinal absorption and are substrates of P-glycoprotein. These features limit the bioavailability of some peptides as orally administered therapeutics. Permeability glycoprotein also known as P-glycoprotein (P-gp; MDR1; ABCB1) is an efflux transporter that is present in the GI tract, BBB, liver, kidneys, and placenta in humans, where it actively transports a wide range of structurally and mechanistically diverse endogenous and xenobiotics across the cell membrane at the energy expense of ATP hydrolysis (Fatoki et al., 2020b). P-gp efflux and CYPs activity can profoundly impact BAPs pharmacokinetics by nutritionally altering their efficacy. The synthetic accessibility score (S) ranges between 1 (easy to make) and 10 (very difficult to make). The partition coefficient (LogP) and solubility coefficient (LogS) contribute to the bioavailability score (Diana et al., 2017; Sanni et al., 2017). Optimization of bioavailability of promising peptides is achievable through side-chain modification or the use of nanodelivery systems. Bioavailability could be affected by food properties including molecular weight, amino acid composition, chain length, sequence of amino acids, net charge, biostability or resistance to hydrolysis by GI tract-based proteases, and hydrophobicity or lipophilicity (Udenigwe et al., 2021; Sun et al., 2020; Wang et al., 2019, 2020). Bioaccessibility of bioactive proteins is vital to the concept of bioavailability, in that bioaccessibility of BAPs is mostly affected by interaction with food matrix constituents such as carbohydrates, lipids, proteins, phenolic compounds, and minerals, which occur during food processing, product formulation, and storage (Sun and Udenigwe, 2020).
The ovalbumin peptides with broad antimicrobial activities (anti-bacterial, antiviral and antifungal) as shown in Table 7, have a common amino acid residue Gly or Pro at position 1 or 2 of the N-terminal. Also, all the peptides with amino acid residue Gly at position 1 of N-terminal showed no anticancer activity, while all the peptides with anti-inflammatory activity have amino acid residue Ile, Gly, Ala, Ser, or Val at position 3 of the N-terminal, and the additional presence of the same amino acid residues at position 2 or 4 improved the anti-inflammatory specificity and sensitivity (Tables s3–s6). Although no antioxidant property was observed for the ovalbumin hydrolysate peptides in this study, the presence of nuclear factor erythroid 2-related factor 2 (NFE2L2) in Figure 2, which is transcription factor that plays a key role in the response to oxidative stress and cell redox homeostasis (https://www.uniprot.org/uniprot/Q16236), showed that certain peptide sequences (VY and QITK) targeting N-formyl peptide receptor 2 (P25090) might possibly contribute to antioxidant property. N-formyl peptide receptor 2 interacts with the scavenger receptor in addition to mediating biological processes, which include positive regulation of superoxide anion generation (https://www.uniprot.org/uniprot/P25090).
| 5. Conclusions | ▴Top |
This study has provided insights on the potential fate of ovalbumin in the GI tract resulting from the combined hydrolytic actions of chymotrypsin, trypsin and pepsin. Data revealed that the GI protease treatment hydrolyzed the allergen sequences present in ovalbumin, which indicate production of a potentially hypoallergenic hydrolysate. Overall, results from binding affinity, pharmacokinetics and physicochemical properties, and therapeutic activity showed that PGF, SSL, GGL, AVL, VY, and IL are promising candidates for further studies. Also, this study reported peptide sequences that possess modulatory effects on existing and potential targets associated with hypertension, diabetes, and immunosuppression. Further work is needed to validate the predicted therapeutic properties of ovalbumin hydrolysate peptides using in vitro and in vivo techniques.
| Supplementary material | ▴Top |
Supplementary materials for this article are available at https://doi.org/10.31665/JFB.2022.17302.
Figure s1. Amino acid sequence of chicken ovalbumin (UniProt ID: P01012).
Figure s2. The cleavage number and sites of each of the proteases, obtained from peptidecutter.
Table s1. Ovalbumin hydrolysate peptides and their SMILES.
Table s2. Predicted binding score of protein-protein interactions obtained from HPEPDOCK server.
Table s3. Anticancer activity prediction.
Table s4. Antimicrobial activity prediction.
Table s5. Anti-inflammatory activity prediction.
Table s6. Antioxidant activity prediction.
Conflict of interest
There are no conflicts of interest to declare.
THF, REA and CCU designed the experiments, THF performed the experiments, analyzed the data, and wrote the draft manuscript. REA and CCU supervised the work and read and edited the manuscript
| References | ▴Top |