| Journal of Food Bioactives, ISSN 2637-8752 print, 2637-8779 online |
| Journal website www.isnff-jfb.com |
Review
Volume 31, September 2025, pages 8-30
Nutritional lipids: an overview of nutritional progress and market trends
Hong Zhang, Xuebing Xu*
Wilmar (Shanghai) Biotechnology Research and Development Center Co. Ltd., Pudong New District, Shanghai 200137, China
*Corresponding author: Xuebing Xu, Wilmar (Shanghai) Biotechnology Research and Development Center Co. Ltd., Pudong New District, Shanghai 200137, China. E-mail: xuxuebing@cn.wilmar-intl.com
DOI: 10.26599/JFB.2025.95031418
Received: August 22, 2025
Revised received & accepted: September 4, 2025
| Abstract | ▴Top |
Nutritional lipids in this text are defined as lipids having nutritional concerns. They are vital components of human diets, delivering essential fatty acids, bioactive compounds, and fat-soluble vitamins that support metabolic health and disease prevention. This review provides an overview of various fatty acids, fatty acid derivatives, and the typical oil-soluble nutrients (unsaponfiable matters) focusing on their nutritional advances and functional benefits. The overview covers all lipid categories existing in nature but with a focus on those having high nutritional concerns and progress. An updated nutritional discussion is given for each group but the latest review papers for the subject have been provided for further reading. An overall picture of nutritional lipids in progress can be expected. For the application channels, functional foods, infant formulas, sport foods, medical foods, and personized nutrition are discussed besides supplementation capsules. The global market is driven by consumers awareness, showing a steady increase. The market distribution is, however, uneven from region to region, indicating that a big market exists for future expansion. There are multiple challenges including oxidation, bioavailability, and regulations, as well as sustainable production technologies. This work integrates scientific insights and industrial trends, offering a scenario for future development.
Keywords: Nutritional Lipids; Bioactive Lipids; Functional Lipids; Fatty Acids; Functional Foods; Structured Lipids; Unsaponifiable Minor Nutrients
| 1. Introduction | ▴Top |
Understanding of lipid nutrition has only gone through fewer than two centuries, although lipids have been used for food for a few thousand years (Gurr et al., 2016). By the mid-1800s, researchers recognized that dietary fats were a concentrated energy source without knowing their metabolic roles. In the 1920s and 1930s, it was discovered that certain fatty acids were essential for growth and health, coining the term essential fatty acids (EFAs). Linoleic acid (ω-6) and later alpha-linolenic acid (ω-3) were identified as crucial dietary components. Around the same time, the fat-soluble vitamins (A, D, E, and K) were identified.
The 1950s–1970s saw growing interest in the relationship between dietary fats and heart disease. Studies found a correlation between saturated fat intake and cardiovascular disease (CVD), leading to dietary guidelines promoting reduced saturated fat consumption. Simultaneously, low-density lipoprotein (LDL) and high-density lipoprotein (HDL) were identified as key players in atherosclerosis. The study further solidified the link between cholesterol and CVD.
In the 1980s–1990s, research revealed the harmful effects of trans fats on LDL cholesterol and heart health. This led to regulatory actions banning trans fats step by step. Conversely, studies on Greenland Inuit populations highlighted the cardiovascular benefits of ω-3 fatty acids, sparking interest in fish oil supplementation.
Recent research has challenged simplistic views on fats, emphasizing the importance of fat quality over quantity. It is suggested that moderate fat intake (including saturated fats) may not be as harmful as previously thought, while ultra-processed fats and refined carbohydrates pose greater risks. Current dietary guidelines (e.g., WHO, USDA, EFSA) now recommend replacing saturated and trans fats with unsaturated fats (especially ω-3s) while recognizing the complex interplay between lipids, metabolism, and chronic diseases. Especially the last decade saw an increasing concern about the nutritional performance of oils and fats, as well as increasing nutritional studies demonstrated by the rising number of research publications.
This progress and the strong perceptions in nutritional lipids have a link with the increase of chronic diseases in the modern time. This seems to have connections with the change of human beings from hunger to overnutrition. In the development of human civilization, we are in a hunger state for most of the time where only in the last 50–100 years we moved into overnutrition step by step (Figure 1) (Livi-Bacci, 2025). Very likely our bodies need more time to adapt to the new situation or make necessary evolution to fit to the new situation. This, however, creates urgency for the mitigation of the increasing problems from governments and public.
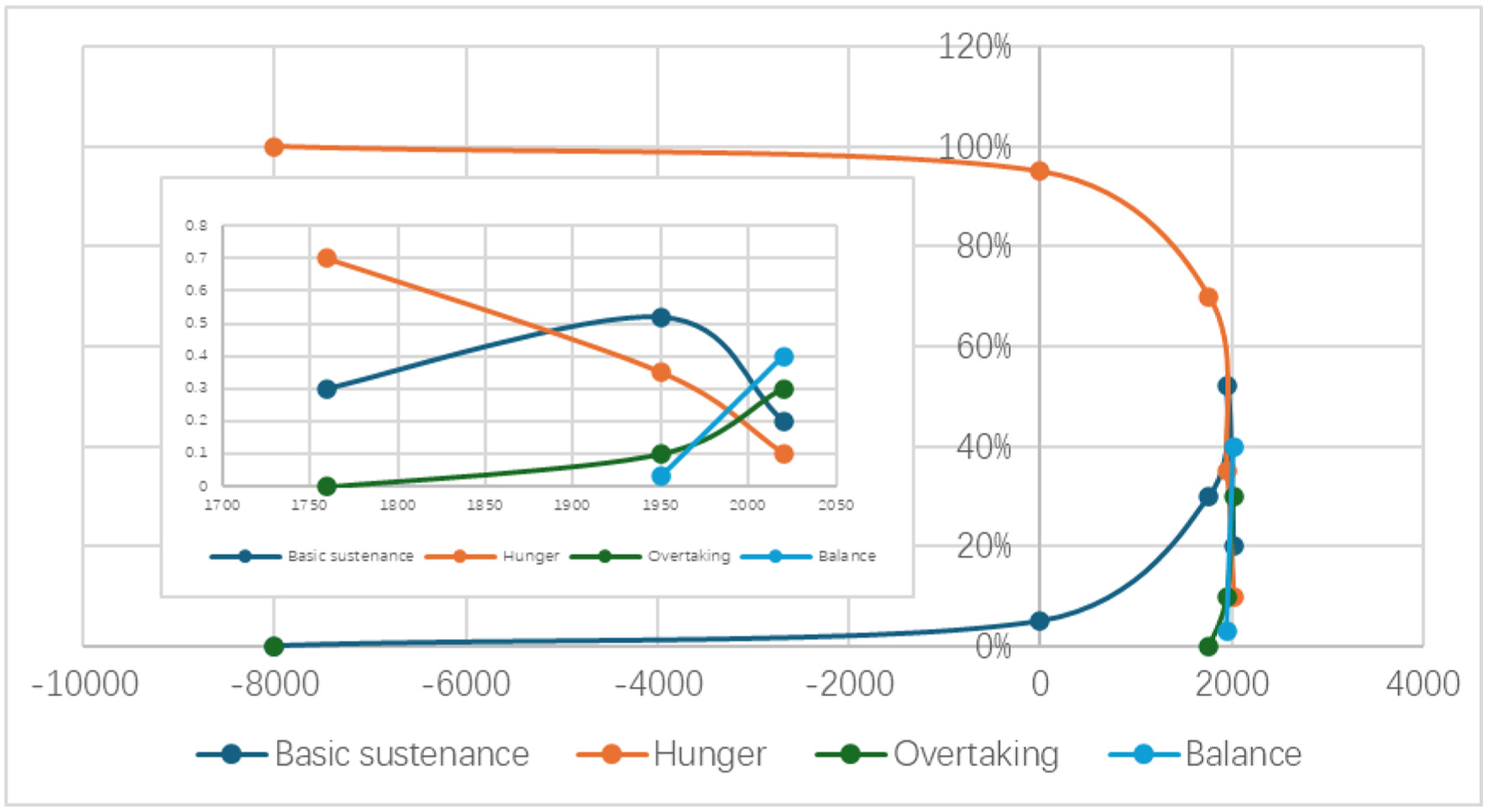 Click for large image | Figure 1. The path for humanity from hunger to overnutrition. |
Lipids are fundamental macronutrients that play a multifaceted role in human health, extending far beyond their traditional role as an energy source. They contribute to cellular structure, serve as carriers for fat-soluble vitamins (A, D, E, and K), and act as precursors to numerous bioactive compounds involved in inflammation, immunity, and metabolic regulation (Chiurchiù and Maccarrone, 2016; David and López-Vales, 2021). Dietary lipids are also crucial for the development and function of the brain and nervous system, particularly in early life stages (Kolko et al., 2023).
There appears a list of terms to describe the nutritional concerns of lipids, such as healthy lipids, nutritional lipids, functional lipids, bioactive lipids, and medical lipids. No official definitions can be found for the terms. Table 1 gives a general understanding of the terms. In this work, nutritional lipids are used to represent all lipids demonstrating a nutritional concern, which cover all the terms.
 Click to view | Table 1. Comparison of lipids in different terms (Gunstone and Herslof, 1992; Gunstone et al., 2007) |
Nutritional lipids represent a diverse and functionally significant class of molecules that extend far beyond their traditional roles as energy sources or structural membrane components (Alhouayek et al., 2021). This review focuses on lipids with demonstrated nutritional and biological significance. These compounds exhibit unique structure-activity relationships where subtle structural variations, such as double bond positioning in PUFAs or headgroup modifications in phospholipids, profoundly influence their bioavailability and metabolic effects (Calder, 2023; Das, 2019a).
The importance of nutritional lipids spans multiple domains of human health and industrial applications. Clinically, these molecules play critical roles in modulating cardiovascular function, cognitive performance, and inflammatory responses (Leuti et al., 2020). Beyond their health benefits, nutritional lipids have become increasingly valuable in industrial applications, with structured lipids engineered for infant nutrition, phytosterols incorporated into functional foods for cholesterol management, and ceramides utilized in advanced skincare formulations. The global market for nutritional lipids is projected to reach $38 billion by 2023 according to the definition in this text, reflecting growing demand across functional foods, pharmaceuticals, and cosmetic sectors (www.strategyr.com). Emerging research continues to reveal novel therapeutic potentials in neurodegeneration, oncology, and metabolic disorders, while lipid-based nanoparticles are revolutionizing drug delivery systems (Stanger and Holinstat, 2023).
Advances in lipid chemistry and biotechnology have enabled the design of lipids with targeted physiological effects (Musso et al., 2018). Technology developments are the strong supports for the supply chain sustainability of nutritional lipids including enzymatic processing of tailored lipids (Xu, 2000; Xu, 2003), lipid production through microorganisms (Nguyen et al., 2025), protection and improved bioavailability by encapsulation (Jala et al., 2025), stabilization of PUFAs (Wang et al., 2021), etc. These developments made nutritional lipids into another level of possibility and high quality. Moreover, food and nutraceutical industries have begun leveraging these lipids to formulate products tailored for diverse health applications, including cardiovascular protection, cognitive enhancement, immune support, and metabolic regulation (Nagao and Yanagita, 2008; Chiurchiù and Maccarrone, 2016). Given the increasing consumer demands for health-promoting ingredients and the rise of personalized nutrition, nutritional lipids represent a strategic frontier for innovation in both dietary and clinical contexts (Das, 2019b).
The objectives of this review are to bridge fundamental and applied research by providing an overall picture of physiological effects of various lipids that receive significant attention from academics; to evaluate industrial situations and critical commercialization challenges; and to identify emerging opportunities in the field. Hopefully the work provides researchers and industry professionals with a current, evidence-based perspective on nutritional lipids.
| 2. Fatty acids: nutritional properties and research advances | ▴Top |
There are more than a few hundred fatty acids reported in nature while only a few dozens are more explored in terms of functions and nutritional understanding. Table 2 is a simple summary of the selected fatty acids which have received more attention. In the following, an insight of fatty acids in categories is discussed with a focus on their nutritional performance.
Click to view | Table 2. Fatty acids and their sources and typical nutritional properties (Gunstone et al., 2007) |
2.1. Short-chain fatty acids (SCFAs)
SCFAs, primarily acetic acid (C2:0), propionic acid (C3:0), and butyric acid (C4:0), are produced through microbial fermentation of dietary fiber in the colon (Tao and Wang, 2025). These metabolites play crucial roles in gut health, metabolic regulation, and immune function (Martindale et al., 2025). Butyrate, in particular, serves as the primary energy source for colonocytes, supporting intestinal barrier integrity and reducing inflammation (Lv et al., 2025). Propionate modulates glucose metabolism by inhibiting hepatic gluconeogenesis, while acetate influences lipid metabolism and appetite regulation (Faradilah et al., 2025). Emerging research highlights SCFAs as key mediators of the gut-brain axis, with butyrate demonstrating neuroprotective effects in preclinical models of Parkinson’s and Alzheimer’s diseases (Cao et al., 2025; Saadh et al., 2025).
Recent advances focus on SCFA-based therapeutics for metabolic and inflammatory disorders (Han et al., 2025). Clinical trials reveal that butyrate supplementation improves insulin sensitivity in obese individuals, and propionate shows promise in reducing visceral fat accumulation (Faradilah et al., 2025). Additionally, SCFAs enhance the efficacy of immunotherapy in colorectal cancer by modulating T-cell activity (Li et al., 2025c). Despite these benefits, challenges remain in optimizing SCFA delivery—oral butyrate is rapidly metabolized in the liver, prompting innovations in colon-targeted formulations (e.g., butyrate-coated nanoparticles, probiotic-engineered strains) (Liang et al., 2025). Future research aims to harness SCFAs for precision nutrition, leveraging microbiome profiling to personalize dietary fiber interventions (Martindale et al., 2025).
2.2. Medium-chain fatty acids (MCFAs, C6:0–C12:0)
MCFAs, comprising caproic (C6:0), caprylic (C8:0), capric (C10:0), and lauric (C12:0) acids, are distinguished by their rapid absorption and metabolism compared to long-chain fatty acids. This phenomenon was discovered by Prof. Babayan in the 60–70s (Bach and Babayan, 1982). Unlike longer-chain fats, MCFAs are transported directly to the liver via the portal vein, where they undergo rapid β-oxidation to produce ketone bodies, making them an efficient energy source (Huang et al., 2021). This unique metabolic pathway has led to their widespread use in medical nutrition therapy, particularly for conditions requiring rapid energy delivery, such as diabetes, obesity, epilepsy, Alzheimer’s disease, metabolic disorders, and athletic performance enhancement (Ameen et al., 2022; Huang et al., 2021). Lauric acid, while technically an MCFA, exhibits metabolic properties intermediate between MCFAs and long-chain fatty acids, with notable antimicrobial and antiviral properties (Jia et al., 2020).
Recent research has highlighted the therapeutic potential of MCFAs in metabolic and neurological disorders (Roopashree et al., 2021). Caprylic acid (C8:0) has gained attention for its role in ketogenic diets, where it enhances ketone production more efficiently than other MCFAs, offering potential benefits for drug-resistant epilepsy and neurodegenerative diseases (Castro et al., 2023). Clinical studies demonstrate that medium chain triacylglycerol (MCT) oil supplementation improves cognitive function in mild cognitive impairment and early Alzheimer’s disease by providing an alternative energy source for glucose-deprived neurons (Dunn et al., 2023). Additionally, MCFAs exhibit anti-obesity effects by increasing energy expenditure and fat oxidation, with capric acid (C10:0) showing promise in reducing visceral adiposity in human trials (Huang et al., 2021).
Emerging applications of MCFAs include infant nutrition and sports performance (Yuan et al., 2022). Structured lipids containing MCFAs are incorporated into infant formulas to mimic the fat composition of human milk, improving fat absorption and digestion in preterm neonates (Jadhav and Annapure, 2023). In sports nutrition, MCT oil is marketed as an ergogenic aid, with evidence suggesting it spares glycogen during endurance exercise (Schönfeld and Wojtczak, 2016). However, challenges remain in optimizing MCFA delivery systems to minimize gastrointestinal distress and improve palatability (Jia et al., 2020). Recent innovations include emulsified MCT formulations and combination products with polyphenols to enhance bioavailability (Roopashree et al., 2021). Future research directions include exploring MCFAs as adjuvants in cancer therapy, where their ketogenic effects may potentiate the efficacy of chemotherapy (Ameen et al., 2022).
2.3. Long-chain saturated fatty acids (LCSFAs)
LCSFAs, including myristic acid (C14:0), palmitic acid (C16:0), and stearic acid (C18:0), have been the subject of extensive research due to their complex roles in human health (Carta et al., 2017). These fatty acids are commonly found in animal fats, dairy products, and tropical oils, and their consumption has been linked to both beneficial and adverse health outcomes (van Rooijen and Mensink, 2020). Recent studies indicate that palmitic acid significantly raises LDL cholesterol levels, increasing CVD risk, while stearic acid appears neutral or even beneficial due to its rapid conversion to oleic acid in vivo (Shen et al., 2025). Myristic acid, though less studied, has been implicated in pro-inflammatory signaling pathways, potentially exacerbating metabolic syndrome (Javid et al., 2020). Emerging evidence suggests that the metabolic effects of LCSFAs may depend on their positional distribution in triglycerides, with sn-2 palmitate showing reduced atherogenicity compared to sn-1/3 variants (Murru et al., 2022).
Recent advances in structured lipid technology have enabled the development of functional fats that mitigate the negative effects of LCSFAs (Salas et al., 2021). Enzymatic interesterification has been used to produce human milk fat substitutes (HMFS) with palmitic acid esterified at the sn-2 position, which improves calcium absorption and reduces fatty stool formation in infants (Annevelink et al., 2023; Xu, 2000). Additionally, stearic acid-rich fats are being explored as cocoa butter alternatives in confectionery, leveraging their unique melting profile and neutral metabolic effects (Shen et al., 2025). In the dairy industry, there is growing interest in modifying bovine milk fat composition through dietary interventions, such as feeding cows oilseeds rich in stearic acid to produce healthier milk fat (van Rooijen and Mensink, 2020). These innovations are supported by advanced analytical techniques, including lipidomics and stable isotope tracing, which provide new insights into the fate of individual LCSFAs in human metabolism (Smith and Bazinet, 2024).
Despite these advances, controversies persist regarding dietary recommendations for LCSFAs (Ceja-Galicia et al., 2025). While traditional guidelines advocate limiting saturated fat intake, newer research emphasizes the importance of considering the food matrix and overall dietary pattern rather than isolated fatty acids (Fatima et al., 2019). For instance, full-fat dairy products, despite their LCSFA content, have been associated with reduced CVD risk in some observational studies, possibly due to the presence of bioactive compounds like phospholipids and odd-chain fatty acids (Murru et al., 2022). Future research directions include investigating the gut microbiota-LCSFA axis, as preliminary evidence suggests that certain LCSFAs may modulate microbial communities involved in metabolic health (Seufert and Napier, 2023). Additionally, there is growing interest in developing plant-based sources of stearic acid through genetic engineering of oilseed crops to create healthier alternatives to traditional saturated fats (Wang et al., 2021).
2.4. Monounsaturated fatty acids (MUFAs)
MUFAs, particularly palmitoleic acid (C16:1ω7), oleic acid (C18:1ω9), and nervonic acid (C24:1ω9), have gained significant attention for their diverse health benefits and metabolic properties. Oleic acid, the most abundant MUFA in the human diet, is a key component of olive oil and is associated with reduced CVD risk through its ability to lower LDL cholesterol while maintaining HDL levels (Santa-María et al., 2023). Recent research highlights its anti-inflammatory effects and its potential role in improving insulin sensitivity in metabolic syndrome (Piccinin et al., 2019). Oleic acid is good also for dermal drug delivery and widely used in cosmetic products (Atef et al., 2022). Palmitoleic acid, a less common but biologically active MUFA, functions as a lipokine that regulates systemic metabolism by enhancing muscle insulin sensitivity and suppressing hepatic lipogenesis (Bermúdez et al., 2022). Emerging evidence suggests it may protect against non-alcoholic fatty liver disease (NAFLD) by reducing hepatic lipid accumulation (Hu et al., 2019). Nervonic acid, a very-long-chain MUFA predominantly found in brain sphingolipids, has shown promise in neurological applications, with recent studies demonstrating its ability to support myelin synthesis and potentially slow progression in multiple sclerosis and neurodegenerative disorders (Namiecinska et al., 2024).
Recent advances in MUFA research have focused on their nutraceutical applications and sustainable production (Desjardins and Rudkowska, 2024). High-oleic oil crops (e.g., genetically modified soybeans and sunflowers) now dominate the food industry due to their oxidative stability and health profile (Wang et al., 2022). Clinical trials with oleic acid-enriched diets demonstrate improved endothelial function and reduced atherosclerotic plaque progression compared to polyunsaturated-rich diets (Tutunchi et al., 2020). Palmitoleic acid is being investigated as a biomarker for metabolic health, with circulating levels inversely correlated with diabetes risk (Bermúdez et al., 2022).
Future research directions include exploring MUFA-epigenetic interactions, particularly oleic acid’s role in DNA methylation patterns linked to longevity (Chu and Yang, 2022) and developing MUFA-producing microbial cell factories through metabolic engineering (Li et al., 2019). Clinical trials are underway to evaluate nervonic acid supplementation in demyelinating disorders, while palmitoleic acid is being tested as an adjunct therapy for NAFLD. Challenges remain in standardizing MUFA biomarkers for clinical use and optimizing delivery systems for nervonic acid’s brain bioavailability (Namiecinska et al., 2024). The growing demand for plant-based MUFA sources is also driving research into algae and yeast production platforms (Phung et al., 2024).
2.5. Omega-6 PUFAs
Omega-6 PUFAs, including linoleic acid (LA, C18:2ω6), γ-linolenic acid (GLA, C18:3ω6), and arachidonic acid (AA, C20:4ω6), are important dietary lipids with distinct metabolic and physiological roles (Jackson et al., 2024). LA, the most abundant ω-6 PUFA in the diet, was first discovered as EFA by Burr and Burr (1929) as discussed in the introduction. The fatty acid serves as a precursor for endogenous synthesis of GLA and AA via elongase and desaturase enzymes (Mercola, 2025). While LA is crucial for maintaining skin barrier function and cellular membrane integrity (Wang et al., 2025), excessive intake, particularly from refined vegetable oils, has been linked to pro-inflammatory eicosanoid production when metabolized into AA (Mercola and D’Adamo, 2023). GLA, found in borage and evening primrose oils, bypasses the rate-limiting Δ6-desaturase step in LA metabolism, making it a promising therapeutic agent for inflammatory conditions like atopic dermatitis and rheumatoid arthritis (Biernacki and Skrzydlewska, 2025). AA, a key component of brain phospholipids, is vital for neurological development, synaptic plasticity, and immune response modulation through its conversion to both pro-inflammatory (e.g., prostaglandin E2) and pro-resolving lipid mediators (e.g., lipoxins) (Crawford et al., 2023).
Recent studies highlight the dual role of ω-6 PUFAs in inflammation, challenging their traditional classification as uniformly pro-inflammatory (Hu et al., 2024). While AA-derived eicosanoids can promote inflammation, its metabolism via 15-lipoxygenase yields lipoxins that actively resolve inflammation (Tredicine et al., 2025). Clinical trials with GLA supplementation demonstrate efficacy in improving symptoms of dry eye disease and menopause-related inflammation, likely due to its conversion to anti-inflammatory dihomo-γ-linolenic acid (DGLA) (Gerges and El-Kadi, 2024). In infant nutrition, AA-fortified formulas show enhanced cognitive outcomes, supporting its critical role in early brain development (Crawford et al., 2023).
Technological advances are enabling precision delivery of ω-6 PUFAs for targeted benefits (Qian et al., 2025). Microencapsulated GLA in functional foods improves stability and bioavailability, while AA-rich algal oils offer sustainable alternatives to animal-derived sources (Ortiz et al., 2024). Research on the gut microbiota-ω-6 axis reveals microbial conversion of LA to conjugated linoleic acid (CLA), which may mitigate metabolic inflammation (Mercola, 2025).
2.6. Omega-3 PUFAs
Omega-3 PUFAs, including α-linolenic acid (ALA, C18:3ω3), eicosapentaenoic acid (EPA, C20:5ω3), and docosahexaenoic acid (DHA, C22:6ω3), are more important nutrients with critical roles in human health as the intake of ω-3 fatty acids is generally insufficient for public (Li et al., 2025a, b). ALA, primarily found in plant sources like flaxseeds and walnuts, was found as EFA a century ago (Burr and Burr, 1929). It serves as a precursor for EPA and DHA, though conversion rates in humans are limited (<10%) due to competition from ω-6 PUFAs for desaturase enzymes (Ghazal and Naffaa, 2025). EPA is a potent modulator of inflammation, primarily through its metabolism into specialized pro-resolving mediators such as resolvins and protectins, which actively resolve chronic inflammation and promote tissue repair (Bodur et al., 2025). DHA, a major structural component of neuronal membranes and retinal tissues, is indispensable for brain development, synaptic function, and visual acuity, with deficiencies linked to cognitive decline and neurodegenerative diseases (Fock and Parnova, 2025). Clinical studies underscore the cardiovascular benefits of EPA and DHA, including reduced triglyceride levels, improved endothelial function, and stabilized atherosclerotic plaques (Amlashi et al., 2025; Zhang et al., 2025a).
Recent research has elucidated novel mechanisms of ω-3 action, particularly in neuroprotection and metabolic health (Li et al., 2025b). Omega-3 fatty acids have a strong link with sleeping (Jackson, 2025) and depression (Kong et al., 2025) as well as for the treatments of bipolar disorder symptoms (Psara et al., 2025). DHA supplementation has been shown to enhance synaptic plasticity and reduce neuroinflammation in Alzheimer’s disease models, with clinical trials demonstrating slowed cognitive decline in early-stage patients (Dyall et al., 2025). EPA’s role in cardiometabolic health has been refined (Tseng et al., 2025). Emerging evidence also highlights ω-3 PUFAs as gut microbiota modulators, where they promote the growth of beneficial bacteria and suppress pro-inflammatory taxa (Jerab et al., 2025). In prenatal nutrition, maternal DHA intake is associated with improved offspring neurodevelopment, including higher IQ scores and reduced risk of attention deficit hyperactivity disorder (Samà et al., 2025).
Technological advances are addressing challenges in ω-3 delivery and sustainability (Marchio et al., 2025). Microencapsulation and emulsion technologies improve the bioavailability of EPA/DHA in functional foods, while algae- and yeast-based production offer scalable, vegan alternatives to fish oil (Jala et al., 2025; Sandoval et al., 2025).
Other ω3 PUFAs such as stearidonic acid (SDA, 18:4 ω3) and docosapentaenoic acid (DPA, 22:5 ω3) are gaining recognition as nutritionally significant ω3 fatty acids, with research highlighting their unique metabolic roles and health benefits. SDA, primarily found in plant sources like Ahiflower oil and echium oil, serves as a more efficient precursor to EPA than ALA, bypassing the rate-limiting Δ6-desaturase step in the omega-3 biosynthesis pathway (Parchuri et al., 2021). This makes SDA-rich oils a promising plant-based strategy for enhancing ω3 status, particularly in individuals with limited fish intake (Baker et al., 2025). Notably, SDA consumption has been linked to anti-inflammatory effects, such as increased interleukin-10 (IL-10) production in humans and reduced pro-inflammatory cytokines in rats, suggesting potential for immune modulation (Parchuri et al., 2021).
DPA, often overshadowed by EPA and DHA, exhibits distinct biological activities and health benefits. It is found in marine sources like fatty fish, seal oil, and, to a lesser extent, in grass-fed meat and certain seaweeds (Drouin et al., 2019; Ghasemi Fard et al., 2021). DPA is not only a metabolic intermediate between EPA and DHA but also has independent roles: it is absorbed into the bloodstream more efficiently than EPA or DHA, incorporates effectively into tissues, and demonstrates potent anti-inflammatory properties (Ghasemi Fard et al., 2021). Research indicates DPA supports cardiovascular health by lowering non-HDL cholesterol and triglycerides, improves cognitive function, and promotes gut microbial diversity (Reinertsen et al., 2024). Importantly, studies in rats and humans show that DPA can retro-convert to EPA and be elongated to DHA, acting as a reservoir for these fatty acids and contributing to their levels in the brain and other tissues (Drouin et al., 2019).
Advances in nutritional science have also clarified their synergistic and individual impacts. For instance, SDA-enriched diets during early life in rats enhanced immune maturation, increasing B-cell function and regulatory T-cells, while DPA’s role in angiogenesis and blood health underscores its importance in vascular function and wound healing (Parchuri et al., 2021; Reinertsen et al., 2024). Furthermore, biomarker studies in humans associate higher circulating DPA levels with reduced mortality risks, emphasizing its clinical relevance (Ghasemi Fard et al., 2021). The development of concentrated SDA and DPA supplements, including sustainable sources like Ahiflower oil and DPA-rich fish oils, offers practical dietary avenues to harness these benefits, particularly for populations with specific nutritional needs or dietary restrictions (Baker et al., 2025; Parchuri et al., 2021). Overall, the growing body of evidence positions SDA and DPA as critical components in the ω3 family, with applications ranging from functional foods to therapeutic interventions for inflammation, metabolic health, and neuroprotection (Drouin et al., 2019; Ghasemi Fard et al., 2021).
2.7. Very long-chain saturated fatty acids (VLSFAs)
VLSFAs, including behenic acid (C22:0), lignoceric acid (C24:0), and cerotic acid (C26:0), are emerging as important but understudied dietary lipids (Lemaitre and King, 2022). These fatty acids, found in trace amounts in plant oils (e.g., peanut, canola) and animal fats, exhibit unique metabolic properties compared to medium- and long-chain saturated fats (Flock and Kris-Etherton, 2013). Unlike their shorter-chain counterparts, VLSFAs are poorly absorbed in the intestine (<30% absorption for C24:0) and appear to have neutral or even beneficial effects on cardiovascular health markers (Lai et al., 2023). Recent studies suggest that dietary VLSFAs may lower LDL cholesterol by reducing intestinal cholesterol absorption, likely through disruption of micelle formation (Lemaitre and King, 2022). In the brain, VLSFAs serve as critical components of sphingolipids in myelin sheaths, with lignoceric acid (C24:0) representing >40% of fatty acids in cerebrosides (Flock and Kris-Etherton, 2013). Emerging evidence indicates that circulating VLSFA levels inversely correlate with incident type 2 diabetes and metabolic syndrome, potentially through modulation of ceramide synthesis pathways (Lai et al., 2023).
Cutting-edge research has unveiled unexpected roles for VLSFAs in neurological and metabolic health (Lemaitre and King, 2022). Lipidomic studies reveal that C24:0-enriched sphingomyelins are associated with reduced risk of Alzheimer’s disease, possibly by maintaining membrane stability and preventing amyloid-β aggregation (Flock and Kris-Etherton, 2013). In X-linked adrenoleukodystrophy, the accumulation of C26:0 due to defective peroxisomal β-oxidation drives neurodegeneration, prompting clinical trials with the oil containing erucic and oleic acids to competitively inhibit VLSFA elongation (Lai et al., 2023). Surprisingly, dietary interventions with high-behenic acid fats (e.g., moringa oil) demonstrate anti-obesity effects in animal models, reducing visceral adiposity (Lemaitre and King, 2022). Advances in analytical techniques like charge-switch derivatization mass spectrometry now enable precise quantification of VLSFAs in biological samples, revealing their tissue-specific distribution and turnover rates (Flock and Kris-Etherton, 2013).
Novel applications are emerging for VLSFAs in functional foods and therapeutics, such as (1) Behenic acid-rich fats are being tested as low-calorie fat substitutes, as their limited absorption provides only 5 kcal/g versus 9 kcal/g for traditional fats (Lai et al., 2023) and (2) C24:0 supplementation is under investigation for demyelinating disorders, with preclinical studies showing enhanced remyelination in multiple sclerosis models (Lemaitre and King, 2022). Key research gaps include elucidating VLSFA-gut microbiota interactions and establishing optimal dietary intake levels (Lai et al., 2023).
2.8. Conjugated fatty acids (CFAs)
CFAs, characterized by their alternating double bonds, include conjugated linoleic acid (CLA, C18:2) and conjugated linolenic acid (CLNA, C18:3), which exhibit unique bioactive properties (Du et al., 2024). CLA isomers, particularly cis-9,trans-11 (rumenic acid) and trans-10,cis-12 CLA, are found in ruminant fats (e.g., dairy, beef) and have demonstrated anti-obesity, anti-inflammatory, and anticancer effects in preclinical models (Badawy et al., 2023). Rumenic acid, the predominant natural CLA isomer, contributes to the health benefits of dairy consumption, including improved lipid metabolism and reduced atherosclerosis risk (Esmaeilnejad et al., 2024). In contrast, trans-10,cis-12 CLA, often used in dietary supplements, reduces adiposity by inhibiting lipoprotein lipase and promoting fatty acid oxidation but may also induce insulin resistance at high doses (Asbaghi et al., 2024). CLNAs, found in pomegranate seed and bitter gourd oils, exhibit even stronger antioxidant and antiproliferative effects than CLA, with α-eleostearic acid (C18:3) showing promise in cancer chemoprevention (George and Ghosh, 2025).
Recent studies have clarified CFA mechanisms and expanded their therapeutic potential (Putera et al., 2023). CLA’s anti-inflammatory effects are now attributed to its modulation of peroxisome proliferator-activated receptor gamma (PPAR-γ) and nuclear factor kappa B (NF-κB) pathways, with clinical trials showing benefits in rheumatoid arthritis at doses of 3–6 g/day (Hoque et al., 2025). In oncology, rumenic acid suppresses tumor growth in breast and colon cancer models by downregulating cyclooxygenase-2 and prostaglandin E2 (González et al., 2025). Advances in microbial biotechnology have enabled high-yield CLA production through fermentation (Badawy et al., 2023). For CLNAs, nanotechnology-based delivery systems (e.g., nanoemulsions) enhance bioavailability, with α-eleostearic acid-loaded nanoparticles showing 3-fold greater tumor uptake in vivo than free fatty acids (Du et al., 2024).
2.9. Odd- and branched-chain fatty acids (OCFAs and BCFAs)
OCFAs, e.g., pentadecanoic acid (C15:0) and heptadecanoic acid (C17:0), and BCFAs, e.g., iso/anteiso-C15:0, iso-C17:0, are emerging as nutritional lipids with unique health implications (Dabrowski and Konopka, 2022). These fatty acids are primarily derived from ruminant fats (dairy, beef) and certain fermented foods, as well as through gut microbial metabolism of dietary fiber (Jenkins et al., 2015). OCFAs have been identified as robust biomarkers of dairy intake and are inversely associated with metabolic disorders, including type 2 diabetes and CVD in large cohort studies (Pfeuffer and Jaudszus, 2016). Mechanistically, OCFAs exhibit mitochondrial modulatory effects, enhancing fatty acid oxidation and reducing oxidative stress, while BCFAs demonstrate anti-inflammatory properties (Dabrowski and Kononka, 2022). Notably, C15:0 has been proposed as an EFA, with studies showing that deficiency leads to mitochondrial dysfunction and cellular senescence in model systems (Jenkins et al., 2015).
Recent advances have elucidated the therapeutic potential of OCFAs and BCFAs in chronic diseases (Pfeuffer and Jaudszus, 2016). C15:0 supplementation (100 mg/day) in preclinical models reversed metabolic syndrome phenotypes, improving insulin sensitivity and reducing hepatic steatosis (Dabrowski and Konopka, 2022). BCFAs, particularly iso-C15:0, show promise in modulating gut microbiota composition (Jenkins et al., 2015). Lipidomic analyses reveal that higher plasma OCFA and BCFA levels correlate with reduced mortality in aging populations, suggesting a role in longevity (Pfeuffer and Jaudszus, 2016). In neuroscience, BCFAs are enriched in myelin membranes, and their dietary intake is linked to improved cognitive performance in elderly cohorts (Dabrowski and Konopka, 2022). Innovations in synthetic biology have enabled the production of OBCFAs, offering sustainable alternatives to animal-derived sources (Jenkins et al., 2015).
2.10. Hydroxy fatty acids (HFAs) and fatty acid esters of hydroxy fatty acids (FAHFAs)
HFAs, such as ricinoleic acid (C18:1-OH) and 12-hydroxystearic acid (C18:0-OH), and FAHFAs, including palmitic acid esters of hydroxystearic acid (PAHSAs), are emerging as bioactive lipids with unique metabolic and anti-inflammatory properties (Wood, 2020). HFAs, found in castor oil and some seed oils, exhibit anti-inflammatory, antimicrobial, and wound-healing effects, with ricinoleic acid demonstrating therapeutic potential in treating inflammatory bowel disease (IBD) (Merheb et al., 2025). FAHFAs, a novel class of endogenous lipids, are particularly notable for their anti-diabetic and insulin-sensitizing effects (Li et al., 2024a). PAHSAs, the most studied FAHFA family, enhance glucose uptake in adipocytes and reduce systemic inflammation, making them promising candidates for metabolic syndrome management (Riecan et al., 2022). Recent studies also suggest that FAHFAs contribute to gut barrier integrity by promoting mucus production and reducing intestinal permeability (Merheb et al., 2025).
Recent research has uncovered the mechanistic pathways and therapeutic potential of HFAs and FAHFAs (Wood, 2020). PAHSAs have been shown to activate the G-protein coupled receptor (GPR120), a key mediator of anti-inflammatory and insulin-sensitizing effects, and to reduce adipose tissue inflammation in obese mouse models (Li et al., 2024a). In humans, lower circulating PAHSA levels correlate with insulin resistance and type 2 diabetes, suggesting their role as biomarkers of metabolic health (Riecan et al., 2022). HFAs like ricinoleic acid are being explored for their antimicrobial properties, particularly against drug-resistant pathogens, due to their ability to disrupt bacterial cell membranes (Merheb et al., 2025). Advances in lipidomics and synthetic chemistry have enabled the identification and large-scale production of FAHFAs, with recent studies demonstrating their stability and bioavailability in functional foods (Wood, 2020).
2.11. Other special fatty acids
There are many more special fatty acids that received attention. Cyclic fatty acids such as sterculic acid (C19:1 cyclopropene) and vernolic acid (C18:1 epoxy) demonstrate potent biological effects, with sterculic acid showing promise in cancer prevention through inhibition of stearoyl-CoA desaturase-1, a key enzyme in tumor lipid metabolism (Kuklev and Dembitsky, 2014). Acetylenic fatty acids like tariric acid (C18:1 acetylenic) exhibit strong antimicrobial and anti-inflammatory activities, particularly against oral pathogens and skin infections (Wang et al., 2024b). Furanoid fatty acids (e.g., furan-C18:2), found in fish and algae, function as endogenous antioxidants, protecting cellular membranes from oxidative damage (Xu et al., 2017). These specialty fatty acids often occur in low concentrations but exert disproportionate biological effects through highly specific mechanisms.
Recent technological advances have enabled detailed study of these rare fatty acids (Elshoura et al., 2024). Lipidomic profiling reveals that furanoid fatty acids accumulate in tissues exposed to oxidative stress, suggesting a protective role that could be harnessed for cardiovascular and aging-related therapies (Xu et al., 2017).
| 3. Fatty acid derivatives: nutritional properties and research advances | ▴Top |
For human consumptions, nutritional lipids are mostly in triacylglycerol forms as in nature. However, many other forms have been developed for various reasons. In the following, nutritional lipids in categories are discussed with the focus on their nutritional properties.
3.1. Natural oils and fats
Nature creates various oils and fats. It is hard to conclude that one is nutritional, and the other is not. Each oil has its characteristics. With proper applications, all can have a good nutritional outcome. However, different oils and fats have different compositions and minor nutrients and give different performances for certain healthy concerns (Zio et al., 2025). Table 3 gives a simple summary of the typical oils and fats selected. The following picks up a few “premier” oils and fats for a bit more discussion.
Click to view | Table 3. Natural oils and fats and their overall nutritional characteristics |
3.1.1. Extra virgin olive oil (EVOO)
EVOO remains the gold standard among edible oils due to its unique composition of monounsaturated fatty acids (74% oleic acid) and bioactive phenolics (e.g., hydroxytyrosol, oleocanthal) (Farhan et al., 2023). Recent research demonstrates EVOO’s superior cardioprotective effects (Lu et al., 2024). A study revealed EVOO phenolics increase gut microbiota diversity, particularly boosting Faecalibacterium prausnitzii abundance, which correlates with improved metabolic health (McNabb et al., 2024). Novel findings indicate hydroxytyrosol glucuronide metabolites cross the blood-brain barrier, potentially explaining EVOO’s neuroprotective effects in Alzheimer’s models (Farhan et al., 2023).
3.1.2. Camellia oil
Camellia oil’s nutritional profile rivals olive oil with higher oleic acid content (82–88%) and greater oxidative stability due to lower PUFA levels (Gao et al., 2024a). Recent studies highlight its unique composition of camelliosides (triterpenoid saponins) that demonstrate potent anti-diabetic effects by inhibiting α-glucosidase and stimulating adiponectin secretion (Qin et al., 2024). The oil’s squalene content (150–300 mg/100g) has been shown to enhance skin barrier function by upregulating filaggrin and loricrin expression in human keratinocytes. Emerging research explores camellia oil’s potential in managing NAFLD.
3.1.3. Linseed/flaxseed oil
As the richest plant source of ALA (50–60%), flaxseed oil continues to demonstrate cardiovascular benefits despite low (<8%) conversion rates to EPA/DHA (Nattagh-Eshtivani et al., 2025). A study showed flaxseed oil reduced arterial stiffness independent of EPA/DHA levels (McNabb et al., 2024). The oil’s lignan content (secoisolariciresinol diglucoside) exhibits unique phytoestrogenic properties, with a recent cohort study associating high flaxseed intake with 31% lower breast cancer risk in postmenopausal women. Innovative microencapsulation techniques now protect the oil’s oxidation-prone ALA while enhancing bioavailability (McNabb et al., 2024).
3.1.4. Walnut oil
Walnut oil’s balanced ω-6/ω-3 ratio (4:1) and high γ-tocopherol content (50 mg/100g) contribute to its emerging role in cardiometabolic health. The oil’s unique polyphenols, particularly ellagitannins, are metabolized by gut microbiota into urolithins that activate nuclear factor erythroid - 2 - related factor 2 (Nrf2) signaling, explaining its anti-inflammatory effects. Recent findings show walnut oil polyphenols inhibit mechanistic target of rapamycin (mTOR) signaling in prostate cancer cells, suggesting potential anticancer applications (Gao et al., 2024b; Zio et al., 2025).
3.1.5. Coconut oil
Virgin coconut oil (VCO) remains controversial but shows unique benefits from its MCTs (50–65%) and lauric acid (C12:0, 45–50%) (Jayawardena et al., 2021). In neurology, VCO increases cerebral ketone bodies (β-hydroxybutyrate +230%) in mild cognitive impairment patients, correlating with improved cognitive scores (Sandupama and Jayasinghe, 2022). Emerging research on fermented VCO shows probiotic-enhanced formulations increase HDL (+12%) without affecting LDL, potentially offering cardiovascular benefits (Jayawardena et al., 2021).
3.1.6. Sea buckthorn oil
This unique oil contains palmitoleic acid (C16:1ω7, 30–40%) rarely found in plants, which shows remarkable skin benefits (Li et al., 2025d). A clinical trial demonstrated sea buckthorn oil improved skin hydration by 28% and reduced transepidermal water loss by 35% in atopic dermatitis patients. The oil’s balanced ω-7/ω-3 ratio (1:1) and tocotrienol content (120 mg/100g) synergistically reduce hepatic steatosis (Chen et al., 2024c). Novel nanoemulsion formulations enhance carotenoid bioavailability 3-fold, expanding therapeutic applications (Chen et al., 2023).
3.1.7. Rice bran oil
Rice bran oil’s γ-oryzanol (1.5–2%) and tocotrienol content contribute to its cholesterol-lowering effects (Sahini and Mutegoa, 2023; Zio et al., 2025). The meta-analysis showed daily consumption reduced LDL by 14% and increased HDL by 8%. The oil’s ferulic acid esters demonstrate potent anti-melanogenic effects, inhibiting tyrosinase activity by 40% compared to kojic acid controls. Emerging applications include nanostructured lipid carriers for targeted cancer therapy, with γ-oryzanol showing selective cytotoxicity against breast cancer cells.
3.1.8. Wheat germ oil
With the highest natural vitamin E content (250–500 mg/100g), wheat germ oil shows unique benefits (Zio et al., 2025). Its octacosanol fraction (50–100 mg/100g) improved athletic performance. The oil’s alkylresorcinols modulate gut microbiota, increasing Akkermansia muciniphila abundance by 3-fold, which correlated with improved glucose tolerance. Supercritical CO2-extracted fractions demonstrate potent anti-leukemic activity.
3.2. Structured triacylglycerols (STs)
There is a stage for scientists to create artificial oils and fats beyond natural oils and fats. One purpose is to create ideal oils and fats for better nutritional functions. The second is to restructure natural oils and fats that have insufficient supplies. STs or structured lipids are the terms of such category. It includes MCTs, medium- and long-chain triglycerides (MLCTs), MLM-type (medium-long-medium, where L refers PUFAs), and OPO/OPL (1,3-dioleoyl-2-palmitoyl glycerol/1-oleoyl-2-palmitoyl-3-linoleoyl glycerol), that represent a class of tailored lipids designed to optimize metabolic and nutritional benefits (Xu, 2000). Table 4 is a summary of STs reported.
Click to view | Table 4. Structured oils and fats and their typical nutritional characteristics |
MCTs (C6-C12) are rapidly absorbed and oxidized, providing quick energy while promoting ketogenesis, making them valuable for managing epilepsy, Alzheimer’s disease, and obesity (Wang et al., 2022). MLCTs, which combine medium- and long-chain fatty acids on the same glycerol backbone, demonstrate enhanced energy metabolism compared to physical MCT-LCT blends, reducing fat accumulation while maintaining EFA supply. MLM-type triglycerides, with PUFAs preferentially esterified at the sn-2 position, can increase the bioavailability and targeted delivery of PUFA for example to the brain (Mu and Porsgaard, 2005). OPO/OPL-structured lipids, specifically designed to resemble human milk fat composition, enhance infant nutrient absorption, cognitive development, and gut microbiota composition (Innis, 2011; Xu et al., 2024). These structured lipids offer distinct advantages over conventional fats by combining the rapid energy delivery of MCFAs with the essential nutritional benefits of PUFAs in optimized molecular configurations (Xu, 2000).
Recent research has significantly advanced our understanding of ST metabolism and applications. Clinical studies demonstrate that MLCTs induce 15–20% greater energy expenditure than traditional LCTs, making them effective for weight management (Wang et al., 2022). OPO/OPL supplementation correlates with sn-2 palmitate reducing calcium soap formation by 40% in preterm infants compared to standard formulas from the optimized lipid matrix (Wei et al., 2024). Technological innovations include enzymatic interesterification using immobilized lipases to produce STs with >90% positional purity, and the development of MLCT-enriched oils from hybrid oilseed crops engineered for specific fatty acid profiles (Xu et al., 2024). Emerging applications include MCT/MLCTs in sports nutrition, where they demonstrate glycogen-sparing effects during endurance exercise, and OPO/OPL in elderly nutrition to combat age-related malabsorption and bone loss (unpublished).
Despite these advances, challenges remain in ST production and application. Scaling up enzymatic processes for commercial ST production requires optimization to maintain cost-effectiveness (Wei et al., 2020; Xu, 2003). Regulatory approval pathways for novel ST formulations vary globally, necessitating harmonized standards (Zou et al., 2019). Future research directions include: 1) precision nutrition approaches tailoring ST compositions to individual metabolic phenotypes using AI-driven lipidomics (Xu et al., 2024); 2) sustainable production through microbial fermentation (Wang et al., 2022); and 3) clinical validation of STs in specialized populations, including metabolic syndrome patients and the elderly (Innis, 2011). The continued development of STs represents a convergence of nutritional science, food technology, and metabolic medicine, offering targeted solutions for diverse health challenges across the lifespan.
3.3. Partial acylglycerols
Partial acylglycerols, particularly diacylglycerols (DAGs), have gained significant attention for their unique metabolic properties and health benefits (Lee et al., 2020). DAG oil, composed predominantly of 1,3-DAG (≥80%), has been extensively studied for its anti-obesity effects, as it reduces postprandial triglyceride levels by 20–30% compared to conventional triacylglycerols (TAGs) due to preferential β-oxidation rather than re-esterification (Ma et al., 2022a). A trial demonstrated that 10 g/day of DAG oil for 12 weeks reduced visceral fat area by 6.5% in obese individuals, attributed to enhanced fat oxidation (Xie et al., 2025). MAGs, particularly monolaurin and monoolein, exhibit antimicrobial and anti-inflammatory properties. MAGs also serve as bioavailable carriers for poorly soluble bioactive compounds, with monoolein-based delivery systems increasing curcumin absorption 5-fold compared to conventional formulations (Zheng et al., 2023).
Recent advances highlight the gut microbiota-modulating effects of partial acylglycerols (Lee et al., 2020). DAG consumption increases Bifidobacterium abundance while reducing Clostridium perfringens, correlating with improved gut barrier function. MAGs, particularly those derived from MCFAs, exhibit prebiotic-like effects. Clinical studies suggest that MAG-enriched diets may alleviate metabolic endotoxemia (Xie et al., 2025).
Despite these benefits, challenges remain in optimizing production technology using enzymes as biocatalysts for the purpose of reducing the production cost as well as composition and stability of partial acylglycerol formulations (Zheng et al., 2023). There are still big spaces for improving to reduce the cost as market prices are relatively high so that the market volume or general public consumption is too low. Furthermore, nutritional esp. clinical studies are still not enough. More nutritional evaluations will be followed also for diversified product profiles and diversified population groups.
3.4. Phospholipids
Phospholipids represent a diverse class of polar lipids that play critical roles in cellular structure, signaling, and metabolism (van der Veen et al., 2017) and cognitive functioning in the whole lifespan (Schverer et al., 2020). Among these, phosphatidylcholine (PC) has emerged as a key nutrient for liver health, with recent studies demonstrating that dietary PC (500 mg/day) reduces hepatic steatosis by 32% in NAFLD patients through enhanced VLDL secretion and PPAR-α activation (He et al., 2025). The choline moiety of PC serves as a methyl donor in one-carbon metabolism. Novel soybean-derived PC fractions enriched in polyunsaturated species (DHA-PC, EPA-PC) demonstrate 3-fold greater bioavailability than triglyceride-forms of ω-3s, with clinical trials showing superior anti-inflammatory effects (Ang et al., 2019; He et al., 2025). Emerging research highlights PC’s role in gut barrier function, where it maintains intestinal mucus integrity by promoting goblet cell differentiation.
Phosphatidylethanolamine (PE) and phosphatidylinositol (PI) exhibit unique metabolic properties (Dong et al., 2023). PE’s role in autophagy has been elucidated through recent studies showing PE/PC ratio determines autophagosome formation efficiency, with PE supplementation improving mitochondrial turnover in aged muscle. PI and its phosphorylated derivatives regulate glucose metabolism. The inositol headgroup of PI shows promise in mental health. Innovative extraction methods now produce marine-derived PE/PI complexes that enhance synaptic vesicle recycling in Alzheimer’s models (Bogojevic et al., 2022).
Phosphatidylserine (PS) and sphingomyelin have gained attention for their neurological benefits (Hussain et al., 2024). Bovine milk-derived PS (100 mg/day) improved cognitive function in mild cognitive impairment patients by enhancing cortical glucose metabolism (Chen et al., 2023). PS’s unique calcium signaling modulation capacity explains its efficacy in stress reduction. Sphingomyelin from milk fat globule membrane demonstrates neurodevelopmental benefits, with infant formula supplemented with 50 mg/L sphingomyelin improving cognitive scores by 5.2 points at 12 months (Singh et al., 2017). The ceramide pathway of sphingomyelin metabolism shows therapeutic potential, with dairy sphingomyelin reducing hepatic ceramide accumulation and improving insulin sensitivity in obese mice (Zhao et al., 2025). Current research focuses on marine phospholipid complexes combining PS, DHA-PC and EPA-PE for synergistic neurological effects (He et al., 2025). More studies are looking into various phospholipids, especially tailor-made phospholipids with enzyme technology (Guo et al., 2005). Table 5 is a summary of such studies.
Click to view | Table 5. Phospholipids and structured phospholipids and their typical functional properties (Ahmad and Xu, 2015; He et al., 2025; Dong et al., 2023) |
3.5. Ether lipids
Ether lipids, characterized by an ether bond at the sn-1 position of the glycerol backbone, comprise two major subclasses: plasmalogens (1-O-alk-1′-enyl-2-acyl lipids) and alkylglycerols (1-O-alkyl-2-acyl lipids) (Magnusson and Haraldsson, 2011). These unique lipids are increasingly recognized for their critical roles in neurological function, membrane dynamics, and oxidative stress resistance (Dorninger et al., 2020). Plasmalogens, which constitute up to 20% of total phospholipids in neural tissues, exhibit neuroprotective properties through their ability to scavenge reactive oxygen species and maintain membrane fluidity (Gomes et al., 2023). Recent clinical studies demonstrate that plasmalogen supplementation (50 mg/day) significantly improves cognitive function in mild cognitive impairment patients (Wang et al., 2024a). The vinyl ether bond of plasmalogens serves as a built-in antioxidant, protecting PUFAs in synaptic membranes from peroxidation, which may explain their therapeutic potential in neurodegenerative diseases such as Alzheimer’s and Parkinson’s (Dorninger et al., 2020).
Alkylglycerols, found abundantly in shark liver oil and human milk, have gained attention for their immune-modulating and anti-inflammatory effects (Wang et al., 2024a). A meta-analysis revealed that alkylglycerol supplementation (100 mg/day) reduced the incidence of upper respiratory tract infections by 40% in elderly populations, likely through enhanced macrophage activity and cytokine regulation (Gomes et al., 2023). These lipids also play a crucial role in gut barrier integrity, with animal studies showing that alkylglycerols upregulate tight junction proteins and reduce intestinal permeability in models of IBD (Magnusson and Haraldsson, 2011). Emerging research highlights the anti-cancer potential of alkylglycerols, particularly in hematological malignancies (Wang et al., 2024a).
Recent advances in nutritional interventions and biomarker discovery have further elucidated the health benefits of ether lipids (Dorninger et al., 2020). Plasmalogen deficiency has been identified as a biomarker for metabolic syndrome, with plasma levels correlating inversely with insulin resistance and hepatic steatosis (Gomes et al., 2023). Dietary strategies to boost plasmalogen levels include consumption of shellfish, organ meats, and certain microalgae, which are rich in ether lipid precursors (Magnusson and Haraldsson, 2011). Krill oil, which contains both plasmalogens and alkylglycerols, improved endothelial function and reduced arterial stiffness in hypertensive patients, outperforming fish oil in vascular health metrics (Wang et al., 2024a). Furthermore, synthetic biology approaches are being explored to produce ether lipids sustainably.
3.6. Fatty acid ethyl esters
Fatty acid ethyl esters (FAEEs), particularly DHA ethyl ester (DHA-EE) and CLA ethyl ester (CLA-EE), represent important bioactive lipid derivatives with distinct metabolic fates and therapeutic applications compared to their triglyceride or free fatty acid forms (Alijani et al., 2025). DHA-EE, the primary component of prescription ω-3 formulations, demonstrates enhanced stability and bioavailability relative to natural triglyceride-bound DHA (Zhang et al., 2019). This improved pharmacokinetic profile stems from preferential hydrolysis by pancreatic lipases and efficient enterocyte uptake, making DHA-EE particularly valuable for treating severe hypertriglyceridemia (Mason et al., 2023). CLA-EE formulations exhibit superior anti-adipogenic effects compared to non-esterified CLA (Alijani et al., 2025). The ethyl ester moiety appears to protect CLA from gastric degradation while facilitating lymphatic absorption, thereby enhancing its bioavailability and tissue distribution (Zhang et al., 2019).
Recent advances have elucidated novel mechanisms of action for these FAEEs (Mason et al., 2023). DHA-EE has been shown to upregulate brain-derived neurotrophic factors in hippocampal neurons more effectively than free DHA, potentially explaining its superior outcomes in mild cognitive impairment trials. Pharmaceutical innovations include self-emulsifying drug delivery systems for DHA-EE that increase oral bioavailability by 200% compared to conventional softgel capsules, and enteric-coated CLA-EE microbeads that prevent gastric isomerization while enabling targeted colonic release for enhanced gut-mediated immunomodulation (Mason et al., 2023).
3.7. Other fatty acid esters
There are many other fatty acid esters which are difficult to go through all one by one. Sugar esters had a story deserving for a discussion. Sucrose polyesters (SPEs), synthetic non-digestible fatty acid esters, have garnered attention as zero-calorie fat substitutes and cholesterol-lowering agents in 90s (Smith et al., 2023). Structurally composed of sucrose esterified with 6–8 long-chain fatty acids, SPEs resist hydrolysis by pancreatic lipase, resulting in minimal intestinal absorption (<0.1%) (Chen et al., 2024b). Originally developed as the active ingredient in Olestra, SPEs have been reformulated to address early gastrointestinal side effects, with newer versions incorporating balanced fatty acid profiles (C12–C18 at 40–60% saturation) to improve tolerability (Anderson and Patel, 2022). Emerging evidence suggests SPEs may act as carriers for fat-soluble vitamins, with nanoemulsified SPE formulations enhancing vitamin D bioavailability by 300% compared to oil-based delivery (Chen et al., 2024b).
Structural modifications, such as medium-chain SPE derivatives (C8–C10), now enable partial hydrolysis (10–15%) to release SCFAs with potential prebiotic effects on gut microbiota (Anderson and Patel, 2022). However, concerns persist about SPEs’ impact on carotenoid absorption, prompting the development of SPE-vitamin A/E-fortified blends that maintain micronutrient status (Smith et al., 2023).
| 4. Unsaponifiable lipids: nutritional properties and research advances | ▴Top |
In natural oils and fats, there are usually less than 1% unsaponifiable compounds, which are bioactive nutrients mostly and give nutritional values and other functional performance such as antioxidative effects. Table 6 gives a collection of the typical compounds. The following is a detailed discussion of the categories.
Click to view | Table 6. Oil-soluble nutrients and their typical functional properties |
4.1. Tocopherols and tocotrienols
The vitamin E family, comprising four tocopherols (α-, β-, γ-, δ-) and four tocotrienols (α-, β-, γ-, δ-), has demonstrated increasingly distinct biological functions beyond their classical antioxidant roles (Atkinson et al., 2008). Recent research highlights the superior neuroprotective capacity of α-tocopherol (Georgousopoulou et al., 2017). However, γ-tocopherol exhibits unique anti-inflammatory properties by scavenging peroxynitrite, with plasma levels inversely correlating with C-reactive protein in metabolic syndrome patients (Sen et al., 2007). The tocotrienols, particularly δ-tocotrienol, show remarkable anti-cancer effects, inducing tumor-selective autophagy through Beclin-1 upregulation while sparing normal cells (Jiang, 2024b). A meta-analysis of seven RCTs confirmed tocotrienol-rich fraction from palm oil reduced liver fat content by 22% in NAFLD patients, surpassing α-tocopherol efficacy (p<0.05) (Phang et al., 2023).
Emerging research reveals isoform-specific transport mechanisms that explain tissue distribution differences (Atkinson et al., 2008). The α-tocopherol transfer protein preferentially binds α-tocopherol with 10-fold greater affinity than γ-tocopherol, while tocotrienols rely on LDL receptor-mediated uptake (Sen et al., 2007). This explains why supplemental γ-tocopherol achieves only 5–10% of α-tocopherol’s plasma levels despite equal dosing. Novel γ-tocopherol/cholate complexes have been developed to bypass this selectivity, increasing γ-tocopherol bioavailability 8-fold in primate models (Jiang, 2024b). In cardiovascular applications, δ-tocotrienol demonstrates unique plaque-stabilizing effects while α-tocopherol shows no benefit in recent trials (Phang et al., 2023).
Current challenges include the pro-oxidant potential of high-dose α-tocopherol (≥800 IU/day), which increased LDL oxidation in 32% of subjects in a pharmacogenomic study (Atkinson et al., 2008). This has spurred development of balanced vitamin E complexes, with a 1:1:1 α-/γ-/δ-tocopherol formulation showing 40% greater antioxidant capacity than α-tocopherol alone in vivo (Sen et al., 2007). Tocotrienol research is advancing through nanoparticle delivery systems, with δ-tocotrienol-loaded nanoemulsions achieving 5-fold higher tumor accumulation versus oral administration in breast cancer models (Jiang, 2024b). Future directions include personalized dosing and combination therapies with ω-3s to enhance membrane incorporation.
4.2. Sterols and sterol esters
Plant sterols (phytosterols) and their esterified derivatives continue to demonstrate potent cholesterol-lowering effects, with recent research refining their mechanisms and expanding applications (Moreau et al., 2018). A meta-analysis confirmed that 2–3 g/day of plant sterol esters reduces LDL cholesterol by 10–12% through competitive inhibition of intestinal cholesterol absorption (Ruscica et al., 2025). Novel microencapsulated sterol formulations now achieve equivalent efficacy at 1.5 g/day by enhancing micellar incorporation (bioavailability increased from <5% to 15%) (Khallouki et al., 2024). Sterol esters derived from pine wood (β-sitosterol oleate) show superior solubility compared to free sterols, enabling incorporation into low-fat foods while maintaining efficacy (Jiang et al., 2024a). Emerging evidence suggests sterol-gut microbiota interactions, with specific bacterial taxa converting sterols into bioactive metabolites that upregulate hepatic LDL receptor expression (Ganamurali et al., 2025).
Beyond cardiovascular benefits, sterol esters exhibit anti-inflammatory and immunomodulatory properties (Moreau et al., 2018). Soy sterol esters have shown promise in metabolic syndrome (Jiang et al., 2024a). Animal studies reveal neuroprotective effects of campesterol palmitate, which crosses the blood-brain barrier to reduce amyloid-β aggregation in Alzheimer’s models (Ganamurali et al., 2025). However, concerns persist about sterol oxidation products, prompting development of antioxidant-stabilized sterol esters using rosemary extract and tocopherol blends.
4.3. Carotenoids
Carotenoids, a class of over 750 naturally occurring pigments, continue to demonstrate diverse health benefits beyond their well-established role as vitamin A precursors (Gholipour-Varnami et al., 2025). Recent advances highlight the differential bioavailability of carotenoid isomers, with all-trans-β-carotene showing 20% greater absorption than 9-cis-β-carotene in an isotope-tracing study (Sereti et al., 2025). Lutein and zeaxanthin, the macular pigments, exhibit neuroprotective effects (Sui et al., 2025). Novel emulsion-based delivery systems have enhanced the bioavailability of lycopene by 300%, overcoming its crystalline insolubility in raw foods (Sereti et al., 2025). Emerging research on apocarotenoids (cleavage products) reveals their role as signaling molecules.
The anti-inflammatory properties of carotenoids are being elucidated through recent mechanistic studies (Sui et al., 2025). Astaxanthin reduced plasma IL-6 by 40% in metabolic syndrome patients by inhibiting NF-κB translocation. Fucoxanthin from brown algae demonstrates unique anti-obesity effects (Sereti et al., 2025). A multi-omics study revealed that β-cryptoxanthin preferentially accumulates in bone tissue, explaining its association with 30% lower osteoporosis risk in epidemiological studies.
4.4. Squalene
Squalene, a triterpenoid hydrocarbon primarily derived from shark liver oil and plant sources (olives, amaranth), has emerged as a compound of significant interest in nutritional research due to its unique antioxidant and lipid-regulating properties (Cheng et al., 2024). Recent studies demonstrate that dietary squalene reduces LDL oxidation by 40% in hypercholesterolemic patients while increasing HDL levels by 15% (Elsharawy and Refat, 2024). A study revealed that olive-derived squalene supplementation significantly improved skin barrier function, increasing stratum corneum hydration by 30% and reducing transepidermal water loss by 25% in atopic dermatitis patients, outperforming conventional emollients (Cheng et al., 2024). The compound’s isoprenoid structure enables unique membrane interactions, with in vitro studies showing squalene incorporation into lipid rafts enhances signal transduction in immune cells (Elsharawy and Refat, 2024). Emerging evidence suggests squalene may serve as a precursor for endogenous CoQ10 synthesis (Cheng et al., 2024).
The chemopreventive potential of squalene has gained attention through recent mechanistic studies (Elsharawy and Refat, 2024). A cohort study associated higher dietary squalene intake (≥300 mg/day) with 35% reduced breast cancer risk (Cheng et al., 2024). In prostate cancer models, squalene nanoparticles (100 nM) induced selective apoptosis in cancer cells, suggesting potential as an adjuvant therapy. However, concerns about oxidative degradation products have prompted development of stabilized squalene formulations using γ-tocopherol and astaxanthin as protective antioxidants.
4.5. Triterpenes
Triterpenes, a structurally diverse class of plant-derived compounds, have gained prominence in nutritional research due to their anti-inflammatory, anticancer, and metabolic regulatory properties (Sharma et al., 2018). A randomized controlled trial demonstrated that boswellic acid (300 mg/day) reduced osteoarthritis pain by 40% (Sen et al., 2024). Novel nanoemulsified triterpene formulations have overcome bioavailability limitations, with β-amyrin nanoemulsions achieving 8-fold higher plasma concentrations than conventional preparations (Sharma et al., 2018). Emerging evidence suggests triterpenes modulate gut microbiota, which correlates with improved gut barrier function in metabolic syndrome patients.
Mechanistic advances have elucidated triterpenes’ epigenetic effects (Sen et al., 2024). Cycloartenol, a key phytosterol precursor, exhibits unique immunomodulatory effects, showing promise in autoimmune disease management (Sharma et al., 2018). The anti-diabetic potential of triterpenes has been refined through the discovery of maslinic acid’s dual agonism on the receptors, achieving 25% greater glucose-lowering than single-target drugs in diabetic mice (Teng et al., 2018).
4.6. Polyphenols
Polyphenols, a diverse class of plant-derived secondary metabolites, continue to demonstrate remarkable multi-target bioactivities that extend beyond their classical antioxidant functions (Zhang et al., 2025b). Recent advances in metabolomics have revealed that polyphenol gut microbiota metabolites (e.g., urolithins from ellagitannins, equol from isoflavones) often exhibit greater bioactivity than their parent compounds, with urolithin A showing 40% greater mitophagy induction in muscle tissue compared to pomegranate extract (Wei et al., 2025). The epigenetic modulation capacity of polyphenols has been elucidated (Suhail et al., 2025). Emerging research highlights polyphenol-mediated gut-vascular barrier protection, where proanthocyanidins from grape seeds reduce bacterial translocation in cirrhosis by strengthening endothelial tight junctions (Wei et al., 2025).
Structure-activity relationships are being refined through cutting-edge research (Zhong et al., 2025). A multi-omics study of 10,000 individuals identified polyphenol-microbiome interactions as key modulators of individual responses (Wei et al., 2025). Clinical applications are expanding, with olive polyphenols (hydroxytyrosol) now in Phase III trials for NAFLD, while theaflavin-rich black tea extracts reduced LDL oxidation by 60% in dyslipidemia patients.
4.7. Oryzanols
Oryzanol, a unique group of ferulic acid esters of phytosterols and triterpene alcohols found in rice bran oil, has emerged as a multifunctional nutraceutical with expanding clinical applications (Dewan et al., 2023). Recent pharmacokinetic studies reveal that γ-oryzanol isomers exhibit differential bioavailability, with cycloartenyl ferulate achieving 3-fold higher plasma concentrations than campesteryl ferulate after oral administration due to superior micellar incorporation (Perez-Ternero et al., 2017). A randomized controlled trial demonstrated that oryzanol-rich rice bran oil (500 mg/day) reduced LDL cholesterol by 18% while increasing HDL by 12% in dyslipidemic patients, outperforming conventional plant sterols in HDL modulation (Kozuka et al., 2013). The antioxidant mechanisms of oryzanol have been elucidated in which its ferulic acid moiety inserts into lipid bilayers, preventing peroxidation chain reactions 40% more effectively than α-tocopherol in model membranes (Perez-Ternero et al., 2017). Emerging research highlights oryzanol’s neuroprotective potential, with 24-week supplementation improving cognitive scores in mild cognitive impairment patients by 2.8 points (Dewan et al., 2023).
The anti-inflammatory properties of oryzanol are being refined through recent mechanistic work (Kozuka et al., 2013). In dermatology, topical oryzanol (2% formulation) improved skin barrier function by upregulating filaggrin and ceramide synthase-3 expression, reducing transepidermal water loss by 35% in atopic dermatitis patients (Dewan et al., 2023). Novel oryzanol-protein complexes with whey or zein have been developed to overcome solubility limitations, increasing oral bioavailability by 8-fold compared to free oryzanol (Kozuka et al., 2013).
4.8. Sesamolins
Sesamolins, a class of unique lignan derivatives primarily found in sesame seeds and oil, have garnered increasing attention for their potent antioxidant and anti-inflammatory properties (Dossou et al., 2023). Recent studies have elucidated that sesamolin and its metabolite sesaminol exhibit differential bioactivities, with sesamolin demonstrating superior Nrf2 activation (3-fold greater than sesaminol) while sesaminol shows stronger NF-κB inhibition (Li et al., 2024b). A study revealed that daily consumption of 50 mg sesamolin-rich sesame oil for 12 weeks reduced oxidative stress markers by 35% and improved endothelial function in hypertensive patients, outperforming conventional sesame oil (Ma et al., 2022a). The gut microbiota-dependent metabolism of sesamolins has been clarified, with specific strains converting sesamolin into bioactive sesaminol triglucoside, which shows 5-fold higher bioavailability than the parent compound (Dossou et al., 2023).
Mechanistic advances have highlighted sesamolins’ multi-target effects in chronic disease prevention. Sesamolin was shown to upregulate LDL receptor expression by 40% in hepatocytes (Ma et al., 2022b). In neuroprotection, sesaminol demonstrated blood-brain barrier permeability and reduced Aβ42 aggregation by 60% in Alzheimer’s models through direct binding to amyloid peptides (Dossou et al., 2023). The anti-cancer potential of sesamolins has been expanded with findings that sesamolin induces ferroptosis in triple-negative breast cancer cells. Novel nanoencapsulation approaches using zein-sesamolin complexes have improved stability against thermal degradation while enhancing oral bioavailability 4-fold.
4.9. Other fat-soluble vitamins
Vitamin A (retinoids and carotenoids) and vitamin K (phylloquinone, menaquinones) have demonstrated expanding roles beyond their classical functions, with recent research uncovering novel mechanisms and clinical applications (Chen et al., 2024a). Vitamin A’s epigenetic regulatory functions have been clarified through studies showing all-trans retinoic acid induces widespread DNA demethylation (Hirota et al., 2025). The gut-liver axis of vitamin A metabolism has been elucidated (Sakwit et al., 2025). Clinically, β-carotene nanoemulsions with enhanced lymphatic targeting achieved 300% higher retinyl ester formation compared to conventional supplements (Chen et al., 2024a). Emerging evidence suggests vitamin A status modulates immunotherapy responses.
Vitamin K research has progressed beyond coagulation to reveal its multiorgan metabolic roles (Sakwit et al., 2025). Long-chain menaquinones (MK-7, MK-9) exhibit tissue-specific accumulation. A multicenter trial demonstrated menaquinone-7 reduced arterial stiffness in hypertensive patients, independent of blood pressure changes (Hirota et al., 2025).
| 5. Existing and emerging product applications of nutritional lipids | ▴Top |
Nutritional lipids have evolved from basic dietary fats to strategic functional ingredients with applications across the food, medical, and nutraceutical sectors (Kupikowska-Stobba et al., 2025). Their role in modulating health outcomes such as inflammation, cognition, metabolism, and cardiovascular function has driven the development of lipid-enriched products designed for specific populations and health goals.
Lipids are primarily consumed through daily foods, making dietary sources critical for ensuring adequate intake of EFAs, phospholipids, and fat-soluble vitamins (Ma et al., 2023). Common food sources include fatty fish, nuts and seeds, dairy products, and plant oils. Fortification of staple foods, such as ω-3-enriched eggs, phytosterol-fortified margarines, and DHA-enhanced dairy products, can improve lipid intake in populations with dietary deficiencies. However, for individuals with specific health conditions, malabsorption issues, or restrictive diets (e.g., veganism), lipid supplements in the form of capsules (e.g., fish oil) or tablets (e.g., vitamin D3) provide a concentrated and bioavailable alternative. Emerging technologies, such as nanoencapsulation and structured lipid formulations, are enhancing the stability and absorption of supplemental lipids, ensuring optimal efficacy (Ma et al., 2023). Balancing dietary sources with targeted supplementation can help meet individual nutritional needs while maximizing health benefits.
5.1. Soft capsules in consumption
Soft capsules are a widely used delivery system for functional lipids such as omega-3s, CLA, and tocotrienols, offering excellent protection against oxidation while ensuring optimal bioavailability (Gullapalli, 2010). The capsule shell is typically composed of gelatin, water, and plasticizers like glycerin or sorbitol to maintain flexibility and prevent brittleness. Gelatin remains the preferred material due to its superior film-forming and oxygen barrier properties, though plant-based alternatives like HPMC are gaining popularity for vegan applications. The lipid fill inside the capsule is carefully formulated with antioxidants (e.g., tocopherols, rosemary extract) and emulsifiers (e.g., lecithin) to enhance stability and absorption. Additional excipients such as MCT oil or phytosterols may be included to improve solubility and functional benefits.
The production of soft capsules primarily relies on rotary die encapsulation, which ensures precise dosing and sealing of the lipid fill within the gelatin or HPMC shell (Gullapalli, 2010). Innovations in encapsulation technology, such as enteric coatings or multilayer lipid systems, further enhance the performance of these capsules by enabling targeted release or improved bioavailability (Jala et al., 2025; Kumar et al., 2024). Challenges such as oxidation control and shell brittleness are addressed through nitrogen flushing, oxygen-barrier coatings, and optimized plasticizer blends. With growing demand for plant-based and allergen-free options, HPMC and pullulan capsules are being refined to match the protective qualities of gelatin. Future advancements may focus on smart capsules with pH-triggered release or synergistic lipid combinations to maximize health benefits (Kumar et al., 2024).
5.2. Functional foods and beverages
One of the most established areas of lipid application is in functional foods, where lipids serve both nutritional and technological roles. Products such as fortified cooking oils, dairy alternatives, nutrition bars, and emulsified beverages often incorporate ω-3 fatty acids, phytosterols, or structured triglycerides to deliver heart and brain health benefits (Kupikowska-Stobba et al., 2025). High-oleic oils are increasingly used due to their oxidative stability and favorable fatty acid profile for cholesterol management (Ma et al., 2023). Additionally, lipid-based delivery of lipophilic vitamins (A, D, E, K) and antioxidants in beverages has been made possible through nanoemulsion and microencapsulation techniques.
5.3. Infant and pediatric nutrition
In infant nutrition, structured lipids such as sn-2 palmitate (OPO) have been developed to mimic the fat composition of human milk, improving fat absorption, calcium uptake, and stool consistency (Ma et al., 2023). PUFAs like DHA and ARA are routinely added to infant formulas to support brain and visual development (de Bhowmick et al., 2023). These advances reflect a convergence of nutritional biochemistry with lipid engineering to deliver health benefits aligned with early-life developmental needs (Chan et al., 2023).
5.4. Medical and clinical nutrition
Medical nutrition products, especially for patients with malabsorption, metabolic disorders, or cachexia, increasingly incorporate tailored lipids (Kiwitz et al., 2025). MCTs are frequently used due to their rapid absorption and metabolism independent of bile salts. Structured lipids with immunomodulatory or anti-inflammatory properties are also being developed for clinical nutrition formulations to support recovery, immune function, and gut health in critically ill or elderly patients (Kiwitz et al., 2025). Parenteral lipid emulsions have evolved to include fish oil-derived ω-3s, offering anti-inflammatory and hepatic-protective effects in enteral and intravenous nutrition.
5.5. Sports and energy products
In sports nutrition, lipids are used for sustained energy release and metabolic support (Cui et al., 2022). MLCTs and emulsified long-chain triglycerides provide both fast-acting and extended energy substrates, improving endurance and fat oxidation capacity during prolonged physical activity. Novel products also incorporate lipids with cognitive and anti-fatigue properties such as PS and CLA, reflecting an intersection between physical and mental performance nutrition (Andrade et al., 2025).
5.6. Personalized and preventive nutrition
As the field of personalized nutrition grows, so does the role of nutritional lipids. Consumers increasingly demand products targeting specific needs, such as metabolic health, cognitive support, or immune enhancement (Kupikowska-Stobba et al., 2025). Custom lipid formulations, such as algal DHA for eye and brain nutrition, CLAs for body control, or DAGs for weight control, are being explored in tailored functional foods and nutraceuticals (de Bhowmick et al., 2023). Preventive applications also include lipid-based carriers for bioactive polyphenols and fat-soluble vitamins that aid in long-term disease risk reduction (Ma et al., 2023).
Overall, the diverse chemical nature and metabolic flexibility of lipids make them uniquely suited for innovation across many food and health product categories. Continued research into structure–function relationships and delivery technologies is expected to expand the application landscape for nutritional lipids in both conventional and personalized formats (Andrade et al., 2025).
| 6. Market situations, challenges, and considerations | ▴Top |
The global market for nutritional lipids is growing steadily, fueled by increasing consumer awareness of their health benefits, scientific advancements, and rising demand for functional foods and dietary supplements. Table 7 gives a collection of typical nutritional lipid products on the global market. Based on the table, a draft calculation of the annual revenue of the listed nutritional lipids is in the range 30–40 billion USD. Figure 2 gives a picture of global distribution by country or region based on a few commercial market reports as well as FAO trade reports. The US takes the lead, and fewer than 10 countries take more than 80% of the share. There could be due to data availability for many countries, but uneven distribution of nutritional lipids should be obvious. The growth in those countries will be expected with the growth of economy. Key segments such as EPA and DHA, MCTs, phospholipids, and plant sterols are driving this expansion, with projections suggesting the market will surpass $10 billion by 2027, growing at an annual rate of around 7.5%.
Click to view | Table 7. Market products of nutritional lipids |
 Click for large image | Figure 2. Global market share of nutritional lipids by country/region. |
Several factors are contributing to this growth. Growing recognition of the role lipids play in cardiovascular, cognitive, and metabolic health is boosting demand, particularly for omega-3s, which are widely used for their anti-inflammatory and brain-supporting properties. At the same time, sustainability concerns are driving interest in plant-based and alternative lipid sources, such as algal oils, insect-derived lipids, and upcycled oils from food processing byproducts. Additionally, personalized nutrition is gaining momentum, with customized lipid formulations becoming increasingly popular.
Regionally, North America and Europe remain the largest markets, supported by strong consumer awareness and strict regulatory frameworks that validate health claims. Meanwhile, the Asia-Pacific region is experiencing the fastest growth, driven by rising incomes, aging populations, and government programs promoting fortified foods. Emerging markets in Latin America and Africa are also seeing increased demand for affordable lipid-fortified products to address nutritional deficiencies.
However, the industry faces challenges, including supply chain sustainability. Regulatory hurdles, such as stringent labeling requirements and approval processes for novel lipid sources, can also slow product development. Furthermore, despite evolving dietary guidelines that emphasize the importance of healthy fats, some consumers remain skeptical about incorporating lipids into their diets.
Looking ahead, technological innovations are expected to shape the future of nutritional lipids. Advances in precision fermentation for single cell lipids, encapsulation techniques to improve bioavailability, and AI-assisted formulation are likely to drive further growth. Strategic collaborations between major food companies and biotech startups are also helping scale sustainable production methods, ensuring that the market continues to evolve in response to global health and environmental needs.
Despite the promising health potential and growing market demand for nutritional lipids, their successful integration into consumer and clinical products remains fraught with challenges. These arise from issues related to stability, sensory quality, scalability, and regulation, which must be carefully addressed to ensure efficacy, safety, and consumer acceptance.
One of the foremost concerns is the oxidative stability of unsaturated lipids, particularly PUFAs like EPA and DHA (Durand et al., 2025). These lipids are highly prone to peroxidation during processing and storage, leading to off-flavors, nutrient loss, and potential formation of harmful compounds (Hennebelle et al., 2024). Encapsulation, use of antioxidants, and packaging under inert atmospheres are common strategies employed to mitigate lipid oxidation, but maintaining stability over shelf life remains a major technical hurdle (Kumar et al., 2024; Wang et al., 2021).
Sensory properties also present challenges. Many health-promoting lipids, especially fish oils, phytosterols, and structured fats, may impart undesirable taste, odor, or mouthfeel, which can reduce consumer acceptance. Formulation technologies such as microencapsulation, emulsification, and the use of masking agents have been developed to overcome these sensory limitations, yet balancing health functionality with palatability continues to be a delicate task (Jala et al., 2025).
From a production and scalability perspective, the synthesis of structured lipids, lipid nanoparticles, or designer emulsions often requires sophisticated processing methods such as enzymatic interesterification, high-pressure homogenization, or microfluidization (Xu, 2003). These processes may increase production costs and complicate quality control, limiting widespread adoption in price-sensitive markets.
Regulatory hurdles are another significant consideration (Durand et al., 2025). Nutritional lipids often fall under functional or novel food categories, requiring rigorous safety assessment and substantiation of health claims (Kumar et al., 2024). While compounds such as DHA, ARA, and phytosterols have secured approval in many regions, newer lipids like OCFAs or structured fats may face uncertain regulatory pathways. Moreover, differences in regional regulatory frameworks add complexity for international product development and marketing.
Individual variability in lipid metabolism also complicates product design and efficacy (Durand et al., 2025). Genetic polymorphisms, age, health status, and gut microbiota composition can influence how lipids are digested, absorbed, and utilized in the body. This calls for more personalized approaches in lipid nutrition and highlights the need for further research into lipid-gene and lipid-microbiome interactions (Kumar et al., 2024).
Environmental and sustainability concerns are increasingly gaining attention. The growing demand for certain lipid sources, such as fish oil and palm oil, raises questions about ecological impact, biodiversity loss, and carbon footprint (Hennebelle et al., 2024). Sustainable sourcing, alternative feedstocks (e.g., algae, microbial oils), and circular production models are essential to ensure that nutritional lipid innovation aligns with global sustainability goals.
Altogether, addressing these challenges is critical to unlocking the full potential of nutritional lipids. Interdisciplinary efforts combining food technology, regulatory science, sensory science, and sustainability principles are essential to drive responsible innovation in this space.
| 7. Conclusions and outlook | ▴Top |
Nutritional lipids represent a dynamic and essential class of food components with extensive implications for human health, food innovation, and global sustainability. Their complexity makes them both a challenge and an opportunity in nutrition science and food technology. As the field advances, understanding how to best classify, deliver, and integrate these lipids into practical applications will be key to maximizing their benefits.
This review has outlined the wide spectrum of nutritional lipids. Each lipid category exhibits unique nutritional functionalities, such as energy provision, anti-inflammatory effects, metabolic modulation, and cognitive support. The diversity of natural sources offers multiple avenues for targeted formulation, yet also demands rigorous evaluation in terms of safety, efficacy, and sustainability.
Improving the nutritional performance of lipids through technological innovation has been a recurring theme. Advances in lipid structuring, encapsulation, and delivery systems have paved the way for better bioavailability and targeted health outcomes. In parallel, consumer demands for clean-label, plant-based, and functional products are driving the development of lipid-containing foods and supplements that align with contemporary health and lifestyle priorities.
Nevertheless, challenges persist. Lipid oxidation, sensory issues, complexity, regulatory barriers, and variable individual responses must be carefully addressed to ensure product efficacy and consumer trust. The shift toward sustainable lipid sources and inclusive supply chains, such as algae oils and byproduct utilization, underscores the broader context in which lipid science now operates.
Looking ahead, strategic directions for future research and industry development include precision lipid nutrition (tailored to genotype and metabolism), functional synergies with other bioactives, and integration with digital health technologies for personalized delivery and monitoring. Investment in interdisciplinary collaboration among lipid chemists, nutritionists, technologists, and policy-makers will be essential to support translational research and scalable innovation.
Ultimately, nutritional lipids stand at the intersection of food, health, and sustainability. Harnessing their full potential will require continued scientific rigor, technological innovation, and ethical foresight, guiding their evolution from basic dietary fats to customized agents of health and well-being.
Acknowledgments
This work received support from Wilmar (Shanghai) Biotechnology Research & Development Center Co., Ltd.
Ethics statement
The work did not involve any ethical issues.
Conflict of interest
The authors declare that they have no conflict of interest.
| References | ▴Top |