| Journal of Food Bioactives, ISSN 2637-8752 print, 2637-8779 online |
| Journal website www.isnff-jfb.com |
Original Research
Volume 23, September 2023, pages 58-67
In Silico Evaluation of Nutri-Pharmacological Potentials of Phytochemicals in Sorghum (Sorghum bicolor) Grains
Toluwase Hezekiah Fatokia, *, Christianah Tosin Baloguna, Oluwafisayomi Taiwo Oluwadarea, Courage Dele Famusiwab, Oyeleke Ridwan Oyebiyia, Blessing Anuoluwapo Ejimadua, Olaolu Ebenezer Lawalb, Busayo Elizabeth Amosunb, Toheeb Olabisi Adeyeyeb, John Adeolu Falodeb
aApplied Bioinformatics Research Laboratory, Department of Biochemistry, Federal University Oye-Ekiti, PMB 373, Oye 371104, Ekiti State, Nigeria
bPhytomedicine and Molecular Toxicology Research Laboratory, Department of Biochemistry, Federal University Oye-Ekiti, PMB 373, Oye 371104, Ekiti State, Nigeria
*Corresponding author: Applied Bioinformatics Research Laboratory, Department of Biochemistry, Federal University Oye-Ekiti, PMB 373, Oye 371104, Ekiti State, Nigeria. E-mail: toluwase.fatoki@fuoye.edu.ng
DOI: 10.31665/JFB.2023.18354
Received: September 17, 2023
Revised received & accepted: September 28, 2023
| Abstract | ▴Top |
Sorghum (Sorghum bicolor L. Moench) ranks as the fifth most widely grown cereal globally, and its grain is gluten-free, containing about 26 phytochemicals which are mostly phenolics. The objective of this study was to predict pharmacokinetics of sorghum grain phytochemicals and some molecular targets that may impact good health in human. The methods used were in silico pharmacokinetic prediction, target prediction, target gene network analysis, docking and molecular dynamics simulation. The results showed that ferulic acid, p-coumaric acid, p-hydroxybenzoic acid, and vanillic acid were blood-brain barrier permeant, while p-coumaric acid and gallic acid have high gastrointestinal absorption. The results indicated that p-coumaric acid has 100% probability of target on aldose reductase and estrogen receptor, while gallic acid has 100% probability of target on alpha-(1,3)-fucosyltransferase 7. The docking analyses revealed that p-coumaric acid bind to aldose reductase with an affinity of −7.759 kcal.mol−1 while gallic acid bind to alpha-(1,3)-fucosyltransferase 7 with binding affinity of −5.512 kcal.mol−1. Overall, binding energy ΔGbind (Total) at 0 ns was slightly higher than that of 100 ns for p-coumaric acid - aldose reductase complex (−56.631 to −53.546 kcal.mol−1). This study provides valuable insights into the potential pharmacological actions of phytochemicals in sorghum grains.
Keywords: Sorghum bicolor; Grain phytochemicals; Computational study; Pharmacokinetics; Molecular targets; Molecular docking and dynamics simulation
| 1. Introduction | ▴Top |
Sorghum (Sorghum bicolor L. Moench) stands as the fifth most cultivated cereal globally and serves as a dietary staple for over 500 million people across 30 countries in semi-arid regions worldwide, including the Americas, Asia, Australia, and Africa (Dahlberg et al., 2011; Fatoki and Sanni, 2019). This gluten-free and drought-tolerant cereal grain plays a versatile role in human nutrition, being used in various forms like bread, malt drinks, beer, and even as livestock feed and a renewable energy source, notably bioethanol (Dahlberg et al., 2011). What makes sorghum unique among major cereal grains is its rich content of bioactive compounds, including phenolic acids, procyanidins, flavonoids, anthocyanins, and the exclusive 3-deoxyanthocyanidins (Yang et al., 2009). These bioactive compounds in sorghum offer numerous health benefits. They support gut microbiota, exhibit a range of biological activities such as anti-inflammatory, antioxidative, anticancer, anti-dyslipidemia, antithrombotic, and antidiabetic properties (Nguyen et al., 2014; Stefoska-Needham et al., 2015; Sanni and Fatoki, 2017; Anunciacao et al., 2018; Ofosu et al., 2021).
While sorghum has traditionally been studied for livestock feed, recent research suggests its potential in human clinical studies (Stefoska-Needham et al., 2015). The nutritional and functional benefits of sorghum which include (i) Sorghum is rich in dietary fiber, which offers several advantages for gut microbiome health, lowers the risk of metabolic diseases, and promotes gastrointestinal well-being (Stefoska-Needham et al., 2015), (ii) Sorghum’s high antioxidant activity has the potential to mitigate oxidative stress, a significant factor in the development of various chronic diseases like diabetes, atherosclerosis, certain cancers, aging, arthritis, and neurological conditions (Awika and Rooney, 2004), (iii) Sorghum also boasts a high content of unsaturated fatty acids in its lipids, which can be beneficial for improving dyslipidemia and thus supporting cardiovascular health (Stefoska-Needham et al., 2015), (iv) Furthermore, the slow digestibility of starch in sorghum has the potential to moderate blood glucose and insulin responses and increase feelings of fullness by reducing the glycemic index of sorghum-based foods (Ezeogu et al., 2005, 2008).
Moreover, it’s crucial to understand that human balanced diets provide essential nutrients necessary for growth, maintenance, and sustainable health. Nutraceuticals, which are biologically active compounds derived from common food sources, enhance this by offering both pharmacological and nutritional benefits, contributing to disease prevention and treatment (Fatoki et al., 2023). Exploring the potential of nutraceuticals in underutilized foods like sorghum could encourage their acceptance and lead to dietary improvements that combat nutrition-related metabolic disorders (Babajide et al., 2013).
The application of computational drug discovery processes such as drug design, optimization, pharmacological and toxicological effect prediction, molecular binding affinity and binding energy prediction, together with genomic mining of novel targets, facilitates translation of natural product such as medicinal plant for clinical use (Harvey et al., 2015; Atanasov et al., 2021; Fatoki et al., 2022).
While it’s been suggested that phytochemicals in sorghum grains could offer protection against diseases in vivo, the full extent of these health benefits remains uncertain. To gain a comprehensive understanding, research should be conducted within a food-health framework that not only explores the impact of individual grain components on physiological processes but also considers the effects of incorporating sorghum-based foods into complete diets. Therefore, the primary objective of this study was to delve into the pharmacokinetics of sorghum grain phytochemicals and analyze their binding interactions with molecular targets that could potentially influence human well-being.
| 2. Methods | ▴Top |
2.1. Ligand preparation
The phytochemical compounds (ligands) in sorghum grains were carefully chosen based on existing literature, which include p-hydroxybenzoic acid, p-coumaric acid, caffeic acid, cinnamic acid, ferulic acid, gallic acid, protocatechuic acid, salicyclic acid, syringic acid, sinapic acid, vanilic acid, apigeninidin, luteolinidin, apigenin, luteolin, naringenin, and trans-resveratrol (Vanamala et al., 2017). The structural information for these ligands was obtained from the PubChem Compound Database (https://pubchem.ncbi.nlm.nih.gov/) in both canonical Simplified Molecular Input Line Entry Specification (SMILES) and Structure Data File (SDF) formats. Conversion of formats was carried out using PyMol for further analysis.
2.2. Pharmacokinetics prediction
In silico screening for ADME (absorption, distribution, metabolism, and excretion) was conducted using the SwissADME server, following the default parameters (Diana et al., 2019).
2.3. Target prediction
The SMILES notations of each ligand were employed for target prediction on the Swiss Target Prediction server (http://www.swisstargetprediction.ch/), with Homo sapiens specified as the target organism (Diana et al., 2019).
2.4. Target gene expression analyses
From the target prediction results, a total of 26 genes were identified for further analysis. These genes include CA2, CA7, CA1, CA6, CA12, CA14, CA9, CA4, CA3, CA5B, CA5A, CA13, AKR1B1, ESR2, MIF, FUT7, ALOX5, MMP9, MM1, MM2, PTPN1, HCAR2, CD38, NOX4, CDK5, and XDH (with their full names available in the results). These gene IDs were then compiled and utilized in various expression network analyses, such as transcription factor enrichment analysis, protein-protein interaction network expansion, and kinase enrichment analysis. This comprehensive analysis was performed using the eXpression2Kinases (X2K) Web server, with the human organism selected as the background reference (Clarke et al., 2018).
2.5. Molecular docking
Molecular docking studies were conducted following the methodology outlined in Fatoki (2023a). To summarize, the crystal structures of the prepared target proteins were subjected to pre-processing in PyMol v2.0.7, involving the removal of water molecules, hetero atoms, and multichain components. Subsequently, both the target proteins and ligands were prepared for docking using AutoDock Tools (ADT) v1.5.6, employing default settings (Morris et al., 2009), and saving the output files in pdbqt format. The actual molecular docking experiments were performed using AutoDock Vina v1.2.3, as described by Trott and Olson (2010) and Eberhardt et al. (2021). Following the docking process, the interactions between the ligands and target proteins were closely analyzed and visualized using the ezLigPlot webserver, accessible at www.dxulab.org/software (Tao et al., 2019). This methodology allowed for a comprehensive examination of the binding interactions between the targets and the ligands of interest.
2.6. Protein-ligand molecular dynamics simulation
Molecular dynamics simulations were conducted for a duration of 100 nanoseconds using Desmond, a software package developed by Schrödinger LLC, as detailed in references (Bowers et al., 2006; Schrödinger, 2018; Fatoki, 2023a). The initial configurations of the protein and ligand complexes for these simulations were derived from the results of the docking studies. Prior to the simulations, the protein-ligand complexes underwent preprocessing using Maestro’s protein preparation wizard. This included optimization and minimization of the complexes. The preparation of all systems was facilitated by the System Builder tool. For the solvent model, an orthorhombic box with TIP3P (Transferable Intermolecular Interaction Potential 3 Points) was selected. The molecular dynamics simulations employed the Optimized Potential for Liquid Simulations (OPLS)-2005 force field, following the methodology outlined by Shivakumar et al. (2010). To mimic physiological conditions, the models were neutralized by adding counter ions in the form of 0.15 M NaCl (Fatoki, 2023b). The simulations were performed in the NPT ensemble, which conserves moles (N), pressure (P), and temperature (T). A temperature of 300 K and a pressure of 1 atm were maintained throughout the simulation. The models were relaxed before the actual simulation process, and trajectory data were saved at regular intervals of 100 picoseconds during the entire simulation period. Post-simulation analysis of the trajectories were analyzed at the end of the simulation to derive the root-mean-square fluctuation (RMSF), root-mean-square deviation (RMSD), and protein-ligand interaction profile. Also, binding free energy was evaluated by using prime molecular mechanics/generalized Born surface area (MMGBSA), based on the following equations:
| 3. Results | ▴Top |
The phytochemical compounds of the sorghum seed grain were investigated. The results of pharmacokinetics prediction in Table 1 presents data indicating that p-coumaric acid, vanillic acid, p-hydroxybenzoic acid, and ferulic acid were predicted to have blood-brain barrier (BBB) permeability. Gallic acid and p-coumaric acid exhibited high gastrointestinal absorption (GIA), with p-coumaric acid having the highest bioavailability score among the two compounds. Additionally, gallic acid has a higher molecular weight compared to p-coumaric acid.
 Click to view | Table 1. Predicted ADME for phytochemical constituents of sorghum grain |
The results of molecular target prediction in Table 2 reveal similarities between p-coumaric acid, gallic acid, and trans-resveratrol in terms of their physicochemical and pharmacokinetic properties. Specifically, these compounds exhibited a 100% probability of targeting several carbonic anhydrases. p-coumaric acid also showed a 100% probability of targeting aldose reductase, estrogen receptor, a 30% probability of targeting macrophage migration inhibitory factor, and a 25% probability of targeting arachidonate 5-lipoxygenase. Gallic acid displayed a 100% probability of targeting several carbonic anhydrases and a 100% probability of targeting Alpha (1–3) fucosyltransferase 7. Finally, trans-resveratrol exhibited a 100% probability of targeting carbonic anhydrase XII.
Click to view | Table 2. Target prediction results for phytochemical constituents of sorghum grain |
In the gene expression network analysis, gene IDs associated with targets having at least a 90% probability were utilized. Figure 1 illustrates the comprehensive interactions among intermediate proteins, kinases (such as CDK1, CDK2, CDK4, ERK1, ERK2, DNAPK, MAPK14, CSNK2A1, and GSK3B), and transcription factors (such as ESR1, KLF4, SUZ12, SMAD4, STAT3, REST, SOX2, RFX5, PPARD, EZH2, CTCF, RELA, RUNX1, RFX5, TP63, MYC, TCF7L2, NANOG and ZBTB33). These interactions are depicted based on their significant hypergeometric (-log10) p-values, providing valuable insights into potential regulatory mechanisms and pathways influenced by the studied bioactive compounds. The results of molecular docking shown in Table 3, showed that the binding affinity of p-coumaric acid, apigenin and luteolinidin to Aldose reductase were −7.759, −10.430 and −10.500 kcal.mol−1 respectively; apigenin binding affinity to xanthine dehydrogenase was −9.196 kcal.mol−1, trans-resveratrol binding affinity to carbon anhydrase XII was −6.328 kcal.mol−1, and gallic acid binding affinity to alpha-(1,3)-fucosyltransferase 7 was −5.512 kcal.mol−1. The interaction of some of the of docking results was presented in Figure 2a–h.
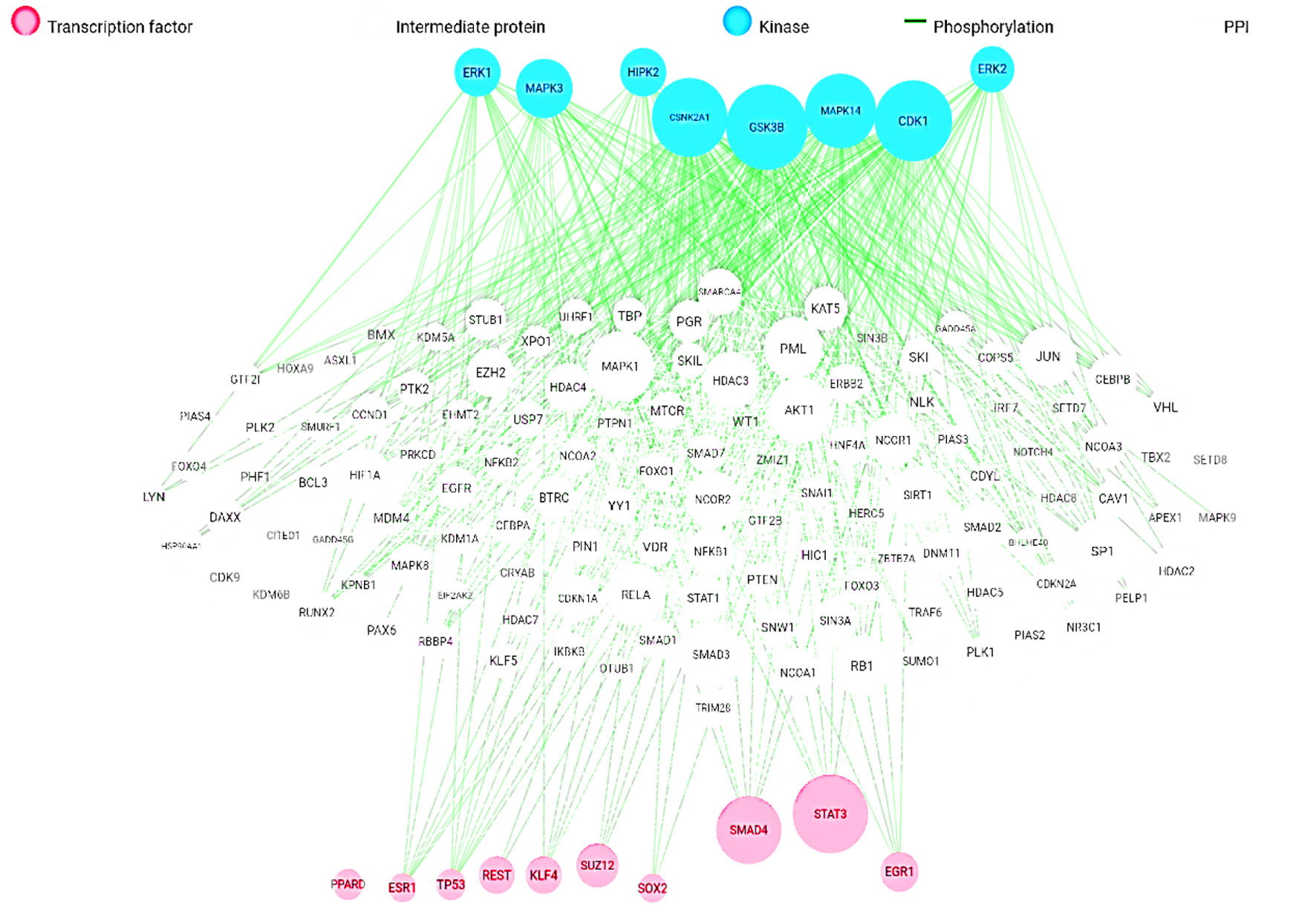 Click for large image | Figure 1. Overall target genes network. |
Click to view | Table 3. Molecular docking results |
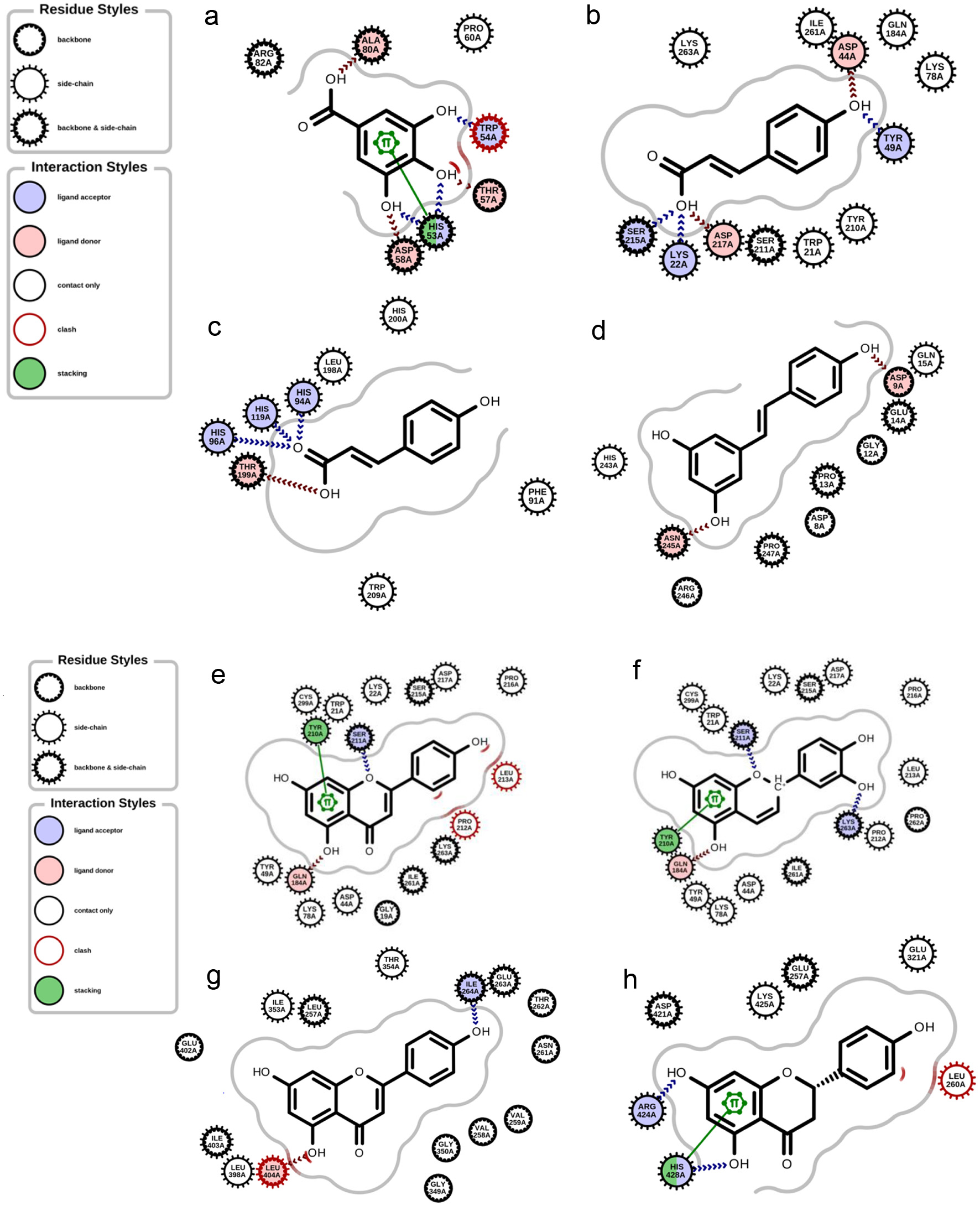 Click for large image | Figure 2. Interaction of the binding poses. (a) gallic acid and alpha-(1,3)-fucosyltransferase 7. (b) p-coumaric acid and aldose reductase. (c) p-coumaric acid and carbonic anhydrase 1. (d) trans-resveratrol and carbonic anhydrase 1. (e) Apigenin and aldose reductase. (f) Luteolinidin and aldose reductase. (g) Apigenin and xanthine dehydrogenase. (h) Naringenin and estrogen receptor |
Molecular dynamic simulations (MDS) were conducted to assess the structural stability of both the protein and the binding status of the ligand in a physiologically relevant environment. Gallic acid and p-coumaric acid were used for MDS analysis based on their high percentage probability of predicted molecular targets and blood-brain barrier (BBB) respectively. The outcomes of these MDS studies have been visualized and are presented in Figure 3a–f, offering valuable insights into the dynamic behavior and interactions of the protein-ligand complexes under realistic conditions. The results of RMSD of p-coumaric acid binding with aldose reductase indicated RMSD of 1.0 - 3.20 Å for the protein at 0 - 100 ns, and 1.00 - 8.00 Å for the ligand at 0 - 100ns (Figure 3d), while gallic acid binding with alpha-(1,3)-fucosyltransferase 7 indicated RMSD of 0.8 - 1.80 Å for the protein at 0 - 100 ns, and 0.6 −4.80 Å for the ligand at 0 - 100ns (Figure 3a). The results of RMSF of aldose reductase showed maximum fluctuate at the center amino acid residues 100–120 and 150–175 (Figure 3e), while RMSF of alpha-(1,3)-fucosyltransferase 7 occurred maximally at the centre amino acid residues 215–230 (Figure 3b). The outcomes of the protein-ligand interactions, including details about the amino acid residues involved in the formation of hydrophobic interactions, hydrogen bonds, water bridges, and ionic interactions, have been illustrated in Figure 3c and f. These figures provide a visual representation of the diverse molecular interactions between the protein and ligand, shedding light on their binding characteristics.
Overall, the results of protein-ligand interactions validated the amino acid residues present in the docking interactions of p-coumaric with aldose reductase and gallic acid with alpha-(1,3)-fucosyltransferase 7 as shown in Figure 2. The binding free energies for all the complexes were computed using MMGBSA at both the 0 ns and 100 ns time points. The obtained results have been presented in Table 4, which provides insights into the stability and energetics of the protein-ligand interactions over the course of the simulation (Figure 3). Overall, binding energy ΔGbind (Total) at 0 ns was higher than that of 100 ns for both the p-coumaric acid - aldose reductase complex (−56.631 to −53.546 kcal.mol−1) and gallic acid - alpha-(1,3)-fucosyltransferase 7 complex (−29.632 to −28.663 kcal.mol−1).
Click to view | Table 4. : Prime MMGBSA binding energy of interaction of p-coumaric acid and aldose reductase and gallic acid with alpha-(1,3)-fucosyltransferase 7, before and after molecular dynamics simulation |
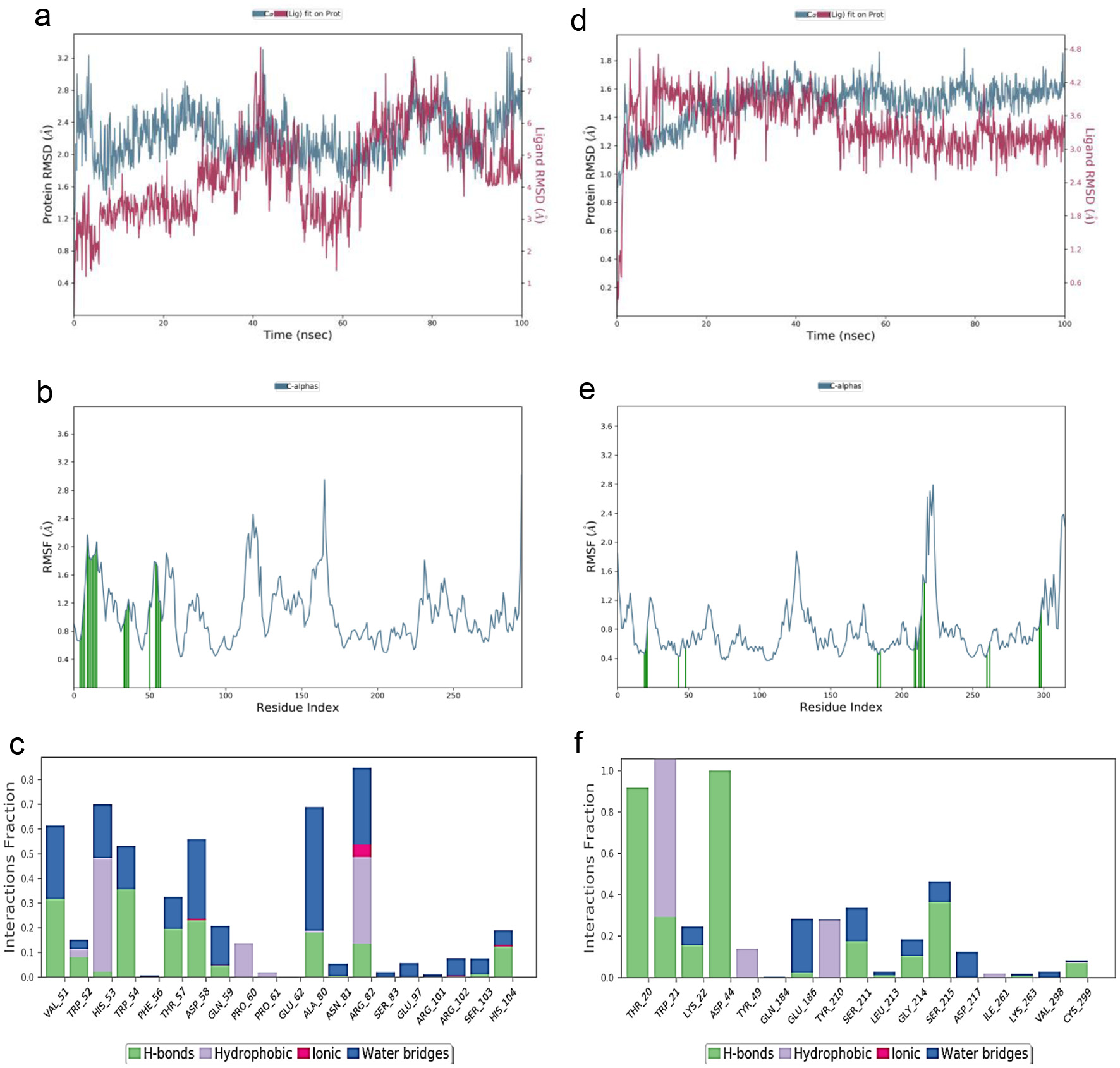 Click for large image | Figure 3. Protein-ligand complex simulation results (a) RMSD of gallic acid and alpha-(1,3)-fucosyltransferase. (b) RMSF of alpha-(1,3)-fucosyltransferase. (c) Interaction profile of the contact between gallic acid and alpha-(1,3)-fucosyltransferase. (d) RMSD of p-coumaric acid and aldose reductase. (e) RMSF of aldose reductase. (f) Interaction profile of the contact between p-coumaric acid and aldose reductase. |
| 4. Discussion | ▴Top |
Among cereals, sorghum has the highest content of phenolic compounds reaching up to 6% (w/w) in some varieties (Awika and Rooney, 2004; Dicko et al., 2006). The phytochemical compounds (phenolic acids, stilbenes, and flavonoids) in sorghum grains includes p-hydroxybenzoic acid, p-coumaric acid, caffeic acid, cinnamic acid, ferulic acid, gallic acid, protocatechuic acid, salicyclic acid, syringic acid, sinapic acid, vanillic acid, apigeninidin, luteolinidin, apigenin, luteolin, naringenin, and trans-resveratrol (Vanamala et al., 2017). The phenolic acids include hydroxybenzoic acid (gallic, syringic, protocatechuic, p-hydroxybenzoic, vanillic, gentistic, and salicylic acids) and hydroxycinnamic acids (caffeic, p-coumaric, ferulic, and sinapic acids), stilbene (resveratrol), flavonoid which include flavanones (e.g. naringenin), and flavones (e.g. luteolin and apigenin) (Li et al., 2023; Sun and Shahrajabian, 2023).
Gallic acid is found in a wide range of natural plants, it is associated with the health of human beings, and it has well-documented anticancer, antibacterial, anti-inflammatory, and antifungal activities, while p-coumaric acid is a plant metabolite with antioxidant and anti-inflammatory properties (Sun and Shahrajabian, 2023). Caffeic acid (3,4-dihydroxycinnamic acid) has been known as a vital source of natural antioxidants in many medicinal plants and has immense use in cancer treatment, while ferulic acid (4-hydroxy-3-methoxycinnamic acid) is a polyphenol that is widely known for its therapeutic potential, showing anti-aging, anti-inflammatory, and neuroprotective effects (Bao et al., 2019; Sun and Shahrajabian, 2023). Resveratrol (3,5,40-trihydroxy-trans-stilbene) is a plant polyphenol, useful in common age-related diseases such as cancer, cardiovascular diseases, type 2 diabetes, and neurological conditions, and it has also positive impacts on metabolism and decrease aging in some organisms (Marques et al., 2009; Sun and Shahrajabian, 2023).
Phenolic compounds had a major contribution to antioxidant activity, and it has been observed that free radical scavenging and antioxidant activity of phenolics (e.g. phenolic acids and flavonoids) mainly depends on the number and position of hydrogen-donating hydroxyl groups on the aromatic ring of the phenolic molecules, and is also affected by other factors, such as H-donating groups (-NH, -SH), and glycosylation of aglycones (Cai et al., 2004). The results of predicted ADME for phytochemical constituents of sorghum grain showed that they are soluble and have high gastrointestinal absorption (GIA), the phenolic acids but not stilbenes and flavonoids, were not affected by p-glycoprotein (p-gp) and some cytochrome p450 (CYP), thus these properties indicate that phenolic acids impact more nutritional benefits in humans that consumes sorghum grains through oral administration due to high bioavailability. Also, the results of this study indicate that only few of the phenolic acids could cross the BBB, and this property may impact positively against neurodegenerative diseases. The concentrations, bioaccessibility, absorption, metabolism and bioavailability of phenolic compounds in food are key factors to optimized in order to maximized their health benefits (Li et al., 2023).
This study revealed that p-coumaric acid, apigenin, and luteolinidin share similar physicochemical and pharmacokinetic properties, along with molecular targets, specifically aldose reductase and the estrogen receptor. Estrogen receptors (alpha and beta) are nuclear transcription factors involved in regulating complex physiological processes in humans. Modulating these receptors with potential therapeutic agents is under consideration for the prevention and treatment of various conditions, including cancer (Ajiboye et al., 2023). Evidence suggests that hormone concentrations and receptor expression may influence the prevalence, progression, and survival of ALS patients (Garcia-Segura et al., 2001; Blasco et al., 2012; de Jong et al., 2013).
Aldose reductase, a member of the aldo-keto reductase superfamily, is an enzyme catalyzing the initial step in the polyol pathway, converting glucose to sorbitol. This enzyme plays a pivotal role in several inflammatory diseases, such as diabetes, cancer, and cardiovascular diseases (Lo et al., 2007; Zeng et al., 2013; Bello et al., 2021). Recent research has shown that aldose reductase is involved in detoxifying aldehydes derived from lipid peroxidation and their glutathione conjugates (Singh et al., 2021). While it’s been identified as a key component of the innate immune response, the precise mechanisms remain unclear (Han et al., 2022).
The target prediction results of this study demonstrate the impact of various phytochemicals found in sorghum grains on several types of carbonic anhydrases (CAs), including I, II, VII, IX, XII, and XIV. These CAs are found both in the cytosol (I, II, and VII) and as membrane-bound forms (IX, XII, and XIV). This suggests that the phytochemicals present in sorghum grains may play a role in mitigating respiratory issues associated with dysregulation of CAs such as amyotrophic lateral sclerosis (ALS) and pneumonia (Fatoki et al., 2022). Also, alpha-(1,3)-fucosyltransferase 7 is an important molecular target for inflammatory bowel disease such as ulcerative colitis, as well as cancer (Barthel et al., 2009; Moriwaki and Moyoshi, 2010).
NADPH oxidase 4 (NOX4) is an enzyme known for producing reactive oxygen species (ROS) and regulating oxidative stress. In infectious disorders, it plays a vital role in defending the host against invading microorganisms (Heyworth et al., 2003). However, in some pathological conditions, NOX4 mediates inflammation and cellular aging through ROS production (Babior, 1999; Wu et al., 2006; Xie et al., 2020). Recent research suggests that NOX4 has a neuroprotective role by regulating ROS and calcium levels, preventing hyperexcitability and neuronal death (Gola et al., 2023). Cyclin-dependent kinase 5 (CDK5) is a protein kinase crucial for neuronal development, synaptic function, and plasticity. It regulates processes like neuronal migration, axonal guidance, and synaptic transmission. CDK5 activity is tightly controlled under normal conditions, but its dysregulation is implicated in neurodegenerative disorders, including ALS (Nguyen and Julien, 2003; Binukumar et al., 2019; Fatoki et al., 2022).
Various bioactive phenolic compounds have been proved to be effective for treatment of diseases, orchestrated through relevant signaling pathways thereby suppress inflammation pathway (e.g., Wnt/β-catenin, phosphatidylinositol3-kinase (PI3K), nuclear factor kappa-B (NF-κB), mitogen-activated protein kinase (MAPK) and NAPDH oxidase) as well as increase antioxidant pathway (e.g., extracellular regulated protein kinases (ERK), nuclear factor erythroid 2-related factor 2 (Nrf2), and endothelial nitric oxide synthase (eNOS)) (Li et al., 2023; Majrashi et al., 2023; Sun and Shahrajabian, 2023).
Moreover, the results of molecular docking showed that the selected phenolic acids acid (p-coumaric acid and gallic acid) have comparable binding affinity to that of stilbene (trans-resveratrol), whereas both phenolic and stilbene have lesser binding affinity than those of tested flavonoids (apigenin, luteolinidin and naringenin). Molecular dynamics (MD) simulations were conducted to assess atomic-level variations in the protein-ligand system and evaluate the stability of the protein-ligand complex in a dynamic environment (Figure 3) (Saini et al., 2021; Fatoki, 2023a). Prime MM-GBSA provided various energy properties, reporting energies for the ligand, receptor, and complex structures, as well as energy differences related to strain and binding. These energy contributions were analyzed in detail (Schrödinger, 2019; Fatoki, 2023a). The root mean square deviation (RMSD) results indicated that both aldose reductase and alpha-(1,3)-fucosyltransferase 7 underwent relatively small conformational changes, suggesting their stability during the simulation (Fatoki, 2023a). The total binding free energy confirmed the stability of the complexes under physiological conditions, indicating their reasonable stability (Figure 3).
| 5. Conclusion | ▴Top |
Sorghum is a globally cultivated cereal grain that is abundant in nutrients, dietary fiber, and bioactive compounds. However, it is often undervalued for human consumption and primarily used as animal feed. This study sheds light on the nutritional potential of sorghum grains, highlighting their richness in beneficial phytochemicals like p-coumaric acid, gallic acid, and trans-resveratrol. These phytochemicals exhibit promising pharmacological profiles against specific target proteins associated with diabetes, cancer, and bowel diseases. Nevertheless, it’s essential to emphasize that further research is necessary to validate the therapeutic effectiveness of these phytochemicals found in sorghum grains, as indicated in this study. This validation can be achieved through in vitro and in vivo experiments, providing more concrete evidence of their potential health benefits.
| References | ▴Top |