| Journal of Food Bioactives, ISSN 2637-8752 print, 2637-8779 online |
| Journal website www.isnff-jfb.com |
Review
Volume 20, December 2022, pages 11-16
Impact of selected novel plant bioactives on improvement of impaired gut barrier function using human primary cell intestinal epithelium
Douglas Bolster*, Lee Chae, Jan-Willem van Klinken, Swati Kalgaonkar
Brightseed, South San Francisco, CA, USA
*Corresponding author: Douglas Bolster, Brightseed, South San Francisco, CA, USA. E-mail: doug.bolster@brightseedbio.com
DOI: 10.31665/JFB.2022.18324
Received: November 6, 2022
Revised received & accepted: December 14, 2022
| Abstract | ▴Top |
Gut barrier function is compromised in the obese state. The N-trans caffeoyltyramine (NCT) and N-trans feruloyltyramine (NFT), two naturally occurring bioactive compounds in hemp hulls, identified using in silico approaches, have the potential to improve gut barrier function and their effects were studied here in vitro. Proliferative human transverse colon epithelial cells were plated and co-cultured with tumor necrosis factor (TNF) along with NCT, NFT or NCT/NFT (2.2 ratio) post-differentiation, over a 48-hour period to induce inflammation and to observe the effects of NCT and NFT. A decrease in transepithelial electrical resistance (TEER) and increase in the intestinal permeability were observed with increased addition of TNF. Co-administration of NCT and NFT demonstrated a dose-dependent and statistically significant reversal of impaired TEER and intestinal permeability. NCT and NFT demonstrated a physiologically relevant reversal of impaired gut barrier function in the setting of inflammation via significant improvement in TEER and percent permeability.
Keywords: Gut barrier function; Bioactives; Gut permeability; N-trans caffeoyltyramine (NCT); N-trans feruloyltyramine (NFT); Hepatic nuclear factor 4α (HNF4α)
| 1. Introduction | ▴Top |
Health promoting properties of food and beverages extend beyond their content of conventional nutrients (vitamins, minerals, dietary fiber, etc.). Naturally occurring bioactive compounds are well recognized for their health promoting properties. Bioactives are small molecules produced by plants, fungi, and microbes that grow in and adapt to environments that present a wide range of challenges to their survival. Bioactives can be widely found in edible plants, herbs, fruits and vegetables (Wallace et al., 2020) and are known to benefit human health. The category of dietary bioactive compounds has been defined by the U.S. National Institute of Health (NIH) and the NIH Office of Dietary Supplements (2018), as “compounds that are constituents in foods and dietary supplements, other than those needed to meet basic human nutritional needs, which are responsible for changes in health status” (National Institutes of Health 2018).
Bioactives such as catechin (found in strawberries), epicatechin (found in dark chocolate), epigallocatechin gallate (found in green tea) have been shown to impart more than just antioxidant properties (Wang et al., 2021; Jean-Marie et al., 2021). These compounds are associated with benefits for heart health, gut health, liver health, skin health, and healthy blood glucose regulation. Not many metabolites have the privilege of crossing the blood brain barrier (BBB), but many bioactive metabolites have been shown to do so and impart neuroprotective benefits (Figueira et al., 2017), thus underscoring how essential they are to our healthy function is important.
The benefits of a Mediterranean diet are widely published, and it is well established that the liver, an essential organ, plays a vital role in maintaining good health. Recent clinical research has shown that consumption of a polyphenol-rich variation of this popular diet is the one factor that makes a noticeable improvement in liver health (Yaskolka Meir et al., 2021). Doubling the daily dose of polyphenols in a standard Mediterranean diet showed a significant reduction in liver fat and inflammation.
The evidence is mounting for an evolved nutrition framework. In this regard, bioactives can inform a new, national standard for dietary guidelines and power an actionable food-as-medicine paradigm for preventative health.
Unlike recommended intakes for vitamins and minerals and dietary fiber, U.S. Dietary Guidelines does not provide recommended intake for bioactives. As a result, it is almost impossible for the general population to understand whether their intake is adequate and if alternate sources should be considered. Although the importance of DHA, EPA, and omega 3 fatty acids is recognized by the U.S. DGA (Wallace et al., 2020), the significance of other bioactives is yet to be highlighted. The U.S. is woefully behind in this regard, as the significance of bioactives is being recognized by nutrition organizations worldwide. The Chinese Nutrition Society, for instance, has published intake recommendations for several bioactives (Wallace et al., 2020).
To further demonstrate the increasing interest and awareness in bioactives, the Academy of Nutrition and Dietetics has published the first dietary recommendation for bioactives, specifically flavan-3-ols, to support cardiometabolic health (Crowe-White et al., 2022). However, flavan-3-ols are only one of many bioactive compounds in edible plants. The published literature recognizes the presence of more than 5,000 known bioactives in plants and food (Wallace et al., 2020) and there are potentially hundreds of thousands more to be discovered. Rapid advances in discovery and validation of bioactives compounds can enable lasting systems reform for the benefit of public health. Artificial Intelligence (A.I.) offers opportunities in nutritional science to discover potential bioactive compounds for known biological targets.
N-trans caffeoyltyramine (NCT, Figure 1) and N-trans feruloyltyramine (NFT, Figure 1) have gained recent attention for their involvement in cellular signaling modulation in several different organs. These two compounds have been identified as bioactive compounds of interest through in silico methods. The methods revealed structural similarities between bioactive compounds and pharmacological approaches for metabolic disease management and their roles as agonists of hepatocyte nuclear factor 4 alpha (HNF4α) (Lee et al., 2021). NCT and NFT were found in a range of edible sources including black pepper and hemp. Hemp, Cannabis sativa, is a well-known plant used for fiber, food, and medicinal purposes. Different subspecies in the Cannabaceae family have varying fruit morphology and levels of psychoactive compounds. When hemp is used as a food source, as in the case of bioactives, the levels of psychoactive compounds such as THC and levels of other compounds such as CBD are at a minimal or undetectable level in the food ingredient (Cerino et al., 2021). Many components of the plant offer value-added function. For example, the hemp hearts (seeds) can be processed for oil and protein, while the hemp hull, the outer coating of the hemp heart, is rich in dietary fiber and also contains protein and fat. Nutritionally, the plant contains the macronutrients carbohydrate, fat, protein, and fiber, along with micronutrients and phytochemicals. The common phytochemicals include phenolic compounds such as flavones, flavonols, and terpenes (Radwan et al., 2021).
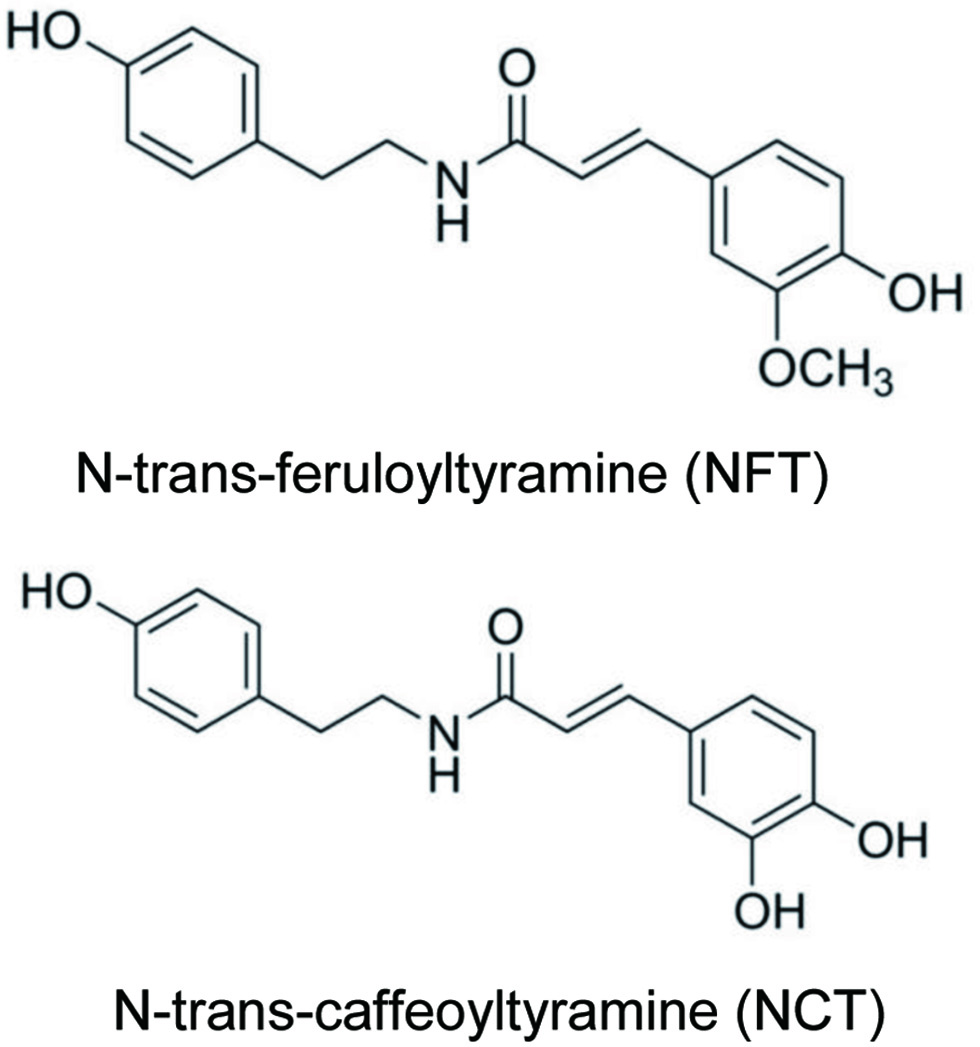 Click for large image | Figure 1. Chemical structures of NFT and NCT. |
HNF4α is a signaling compound, known as a nuclear transcription factor, that plays a role in regulating metabolism in the liver. Within the liver, HNF4α regulates gluconeogenesis and lipid metabolism, more specifically hepatic lipid storage (Lee et al., 2021). It has also been identified in the intestine, pancreas and kidney where it serves other functions. Within the intestine, HNF4α influences intestinal permeability and is associated with inflammatory bowel diseases, although its role is unclear. Agonists of HNF4α activate this transcription factor. Synthetic agonists such as alverine for inflammatory bowel disease (IBD) management and benfluorex, formerly approved for type 2 diabetes management, are pharmaceutical approaches to enhance HNF4α.
Beyond the liver, HNF4α has been documented to exert numerous effects, specifically on gut barrier and intestinal function (Babeu and Boudreau 2014). Specifically, HNF4α impacts mucin production, which is a key component of the gut barrier. It also regulates expression of tight junction proteins and Paneth cell differentiation. Both of these aspects influence the human host’s ability to maintain homeostasis with the gut microbiota, and thus influence both gut health and overall human health.
Molecules that are structurally similar to known HNF4α agonists can also enhance HNF4α activity. Two such naturally occurring compounds found in plant cell walls are NCT and NFT (Figure 1). The direct interaction of NCT and NFT with HNF4α makes NCT and NFT potent agonists of HNF4α. NCT is the precursor to NFT, with a reaction mediated by caffeoyltyramine-O-methyltransferase, which replaces the methyl group in NCT with a hydroxyl group to form NFT.
These two compounds (Figure 1) have similar agonist effects on HNF4α; however, NCT appears to have a greater potency on insulin and HNF4α mRNA expression than NFT (Lee et al., 2021). In cell culture, both NCT and NFT reduced fat storage in both qualitative and quantitative assays. Furthermore, mechanistic investigation of NCT’s effect shows a linkage between the gene regulation and inflammatory bowel diseases (Lee et al., 2022). NCT specifically upregulates genes that influence nitric oxide synthesis (plays a complex role in IBD development), sodium/bile acid transporters (relevant in Crohn’s disease), short-chain fatty acid transporters (relevant to ulcerative colitis), dual oxidase 2 which generates reactive oxygen species (ROS, relevant in early IBD development) among several others. These genes are downregulated in the diseased state. While the aforementioned data have been generated in pre-clinical studies, it shows proof of concept and provides the basis for further clinical investigation of NCT and gut health. In cell culture, both NCT and NFT reduced fat storage in both qualitative and quantitative assays.
The phenomena noted in cell culture above have been reproduced in obese mice upon administeration of NCT for 10 weeks (Veeriah et al., 2022). What is notable about this study, is that the mice were administered NCT orally, which simulates dietary intake of NCT. This models the way that NCT would be handled when consumed from a natural source. NCT resulted in 30–40% weight gain reduction, in spite of no differences in energy intake and no differences in stool fat excretion, which would indicate fat malabsorption. Moreover, these mice were fed a high fat diet to induce obesity, and the reduced weight gain indicated that NCT could mediate the effects of the diet. The remaining explanation for the differences in body weight and fat mass was due to increased fat oxidation when NCT was consumed. Indeed, NCT increased fatty acid oxidation via a secondary mechanism, mediated by changes in mitochondrial function. Markers of mitochondrial stress were greatly reduced by NCT, which suggests that NCT could play a role in slowing the progression of nonalcoholic fatty liver disease (NAFLD) to nonalcoholic steatohepatitis (NASH). Another indicator of NAFLD progression to NASH is inflammation. Interleukin-6 (IL-6) and TNF were reduced in the mice consuming NCT. These findings were also confirmed in cultured human hepatocytes.
The use of in silico approaches enabled the identification of NCT and NFT as potential bioactive compounds to affect several aspects of human health. Previous in vitro and pre-clinical studies demonstrated that NCT and NFT impact hepatic homeostasis as well as gut barrier function. The HNF4α agonist effects of NCT and NFT suggest that they could exert protective effects on gut barrier function, particularly in an inflammatory environment. Although HNF4α function was not assessed in this current research endeavor, the mechanistic foundation previously published suggests that the roles of NCT and NFT in this respect are via an HNF4α-mediated function (Babeu and Boudreau 2014). The aim of this study was to determine the effects of hemp-derived NCT and NFT on rescuing impaired gut barrier function using human stem cell derived colon cells, to further understand the role of NCT and NFT in gut health.
| 2. Materials and methods | ▴Top |
2.1. Bioactive materials
NCT and NFT were obtained from Sundia MediTech Company (Shanghai, China) and suspended in dimethyl sulfoxide (DMSO, Sigma, St. Louis, MO, USA). TNF (R&D Systems, Minneapolis, MN, USA) was prepared according to manufacturer’s instructions.
2.2. Cell culture and compound treatment
Proliferative human transverse colon epithelial cells were plated using protocols established by the contract research organization into 4 × 96 well RepliGut® (Altis Biosystems, Durham, NC, USA) transwell plates coated with a thin layer of hydrogel. Cultures were monitored daily using a bright field microscope, and on day 4 the transverse colon epithelial monolayers become confluent. At this time, the media were changed from expansion media (Altis) to differentiation media (Altis). The cells were then cultured for an additional two days. On day seven, NCT, NFT or NCT/NFT (2.2 ratio) along with TNF, to induce inflammation, were diluted in differentiation media (30 ng/mL) and added to the apical and basal compartment of the transwell cultures at t = 0. All selected conditions were repeated 3 times. Cells were monitored for 48 hours, with readings taken at 0, 24, 45 and 48 hours.
2.3. Transepithelial electrical resistance (TEER)
Transepithelial electrical resistance was measured in all transwell cultures at t = 0 hour prior to the addition of compounds to determine the baseline TEER using an Epithelial Volt/Ohm Meter (World Precision Instruments, EVOM2, Sarasota, FL, USA) and STX100C96 Electrode. TEER was also measured at t = 24 hours, t = 45 hours, and t = 48 hours. The surface area of a 96-well transwell is 0.143 cm2 per well. Corrected TEER was calculated using the formula below.
2.4. Supernatant collection
After the TEER time point at t = 48 hours, apical and basal supernatants were transferred to a 96-well polypropylene storage microplate (Thermo Fisher, Raritan, NJ, USA), sealed with Sterile AlumaSeal II Sealing Film (Genesee, El Cajon, CA, USA), and stored at −80 °C.
2.5. Permeability assay
After collection of supernatants, a permeability assay was run on all transwells. Forty-five hours after the addition of compounds, 75 μL of 0.5 mg/mL 40 kD M.W. Fluorescein labeled dextran (FITC-dextran, Sigma) solution made in differentiation media was placed in the apical compartment of each transwell and 210 μL of differentiation media was placed in the basal compartment of each transwell. Transwell cultures were incubated for 3 hours at 37 °C with 5% carbon dioxide to allow for the transfer of FITC-dextran into the basal compartment. Two aliquots of 90 μL of media were collected from the basal compartment of each transwell and fluorescence was measured (Ex: 485, Em: 535) for each sample (n = 2 samples/well) using a Synergy H1 Plate Reader (Aglient, Santa Clara, CA, USA).
“Raw fluorescence (RF) of media” was obtained by measuring differentiation media without FITC-Dextran to determine background fluorescence of the media. “RF of no cell control,” was obtained from basal compartment media of hydrogel-coated transwells that were not seeded with epithelial cells to normalize permeability measurements. “Relative Permeability (% of No Cell Control)” was calculated using the formula below.
Any remaining solution was gently aspirated, and cells were fixed on transwells with methanol. After three 5-minute rinses with phosphate buffered saline (PBS), transwells were stored in PBS at 4 °C.
2.6. Statistical analysis
Statistical analysis was performed using GraphPad Prism software version 9.4.1 (GraphPad Inc., La Jolla, CA, USA). Two-way ANOVA of means from n = 3 samples per group was performed to measure simple effects within treatment groups. Tukey’s range test was performed to correct for multiple comparisons. Differences between the groups were considered significant at p < 0.05.
| 3. Results and discussion | ▴Top |
Administration of NCT improved the change in TEER as a result of TNF administration in a dose-dependent manner after 48 hours, with increasing improvements from 10 to 20 to 40 μM (Figure 2a). Values shown are relative to t = 0. As little as 10 μM NCT resulted in improvements in % permeability that nearly countered the effects of TNF administration (Figure 2b). Higher doses of NCT (20 μM and 40 μM) exhibited a similar improvement to the 10 μM NCT dose.
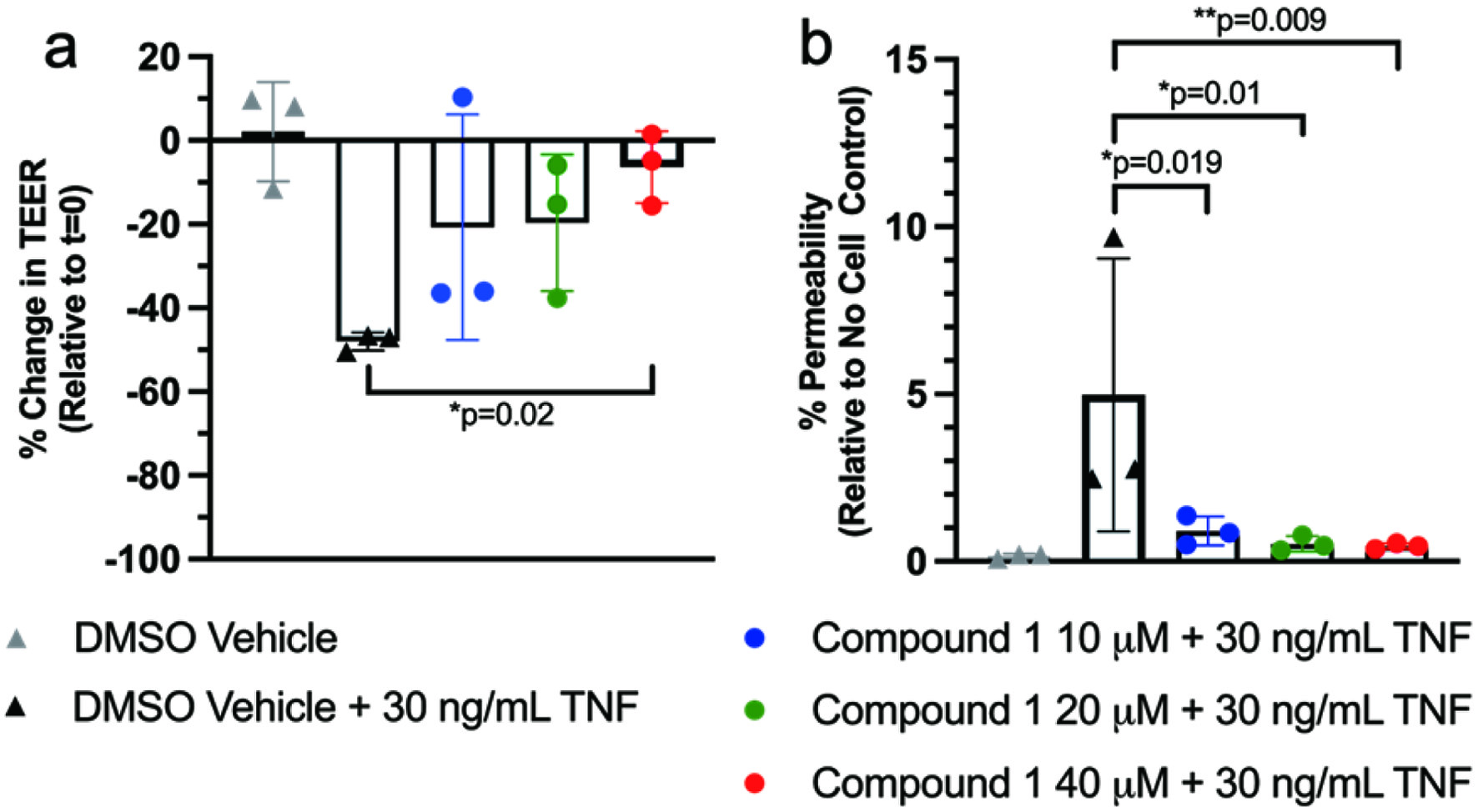 Click for large image | Figure 2. Administration of NCT protected against (a) decreased TEER and (b) increased permeability with TNF-induced inflammation. Compound 1 refers to NCT. |
Administration of NFT improved TEER at 10 μM; however, increasing the concentration of NFT to 20 and 40 μM did not show the same dose-dependent effect that NCT exerted (Figure 3a). Similarly, the lowest dose of NFT improved % permeability to the extent that the effects of TNF were nearly completely mitigated (Figure 3b). As the dose of NFT increased, so did the variability of the % permeability such that a dose-dependent effect was not observed.
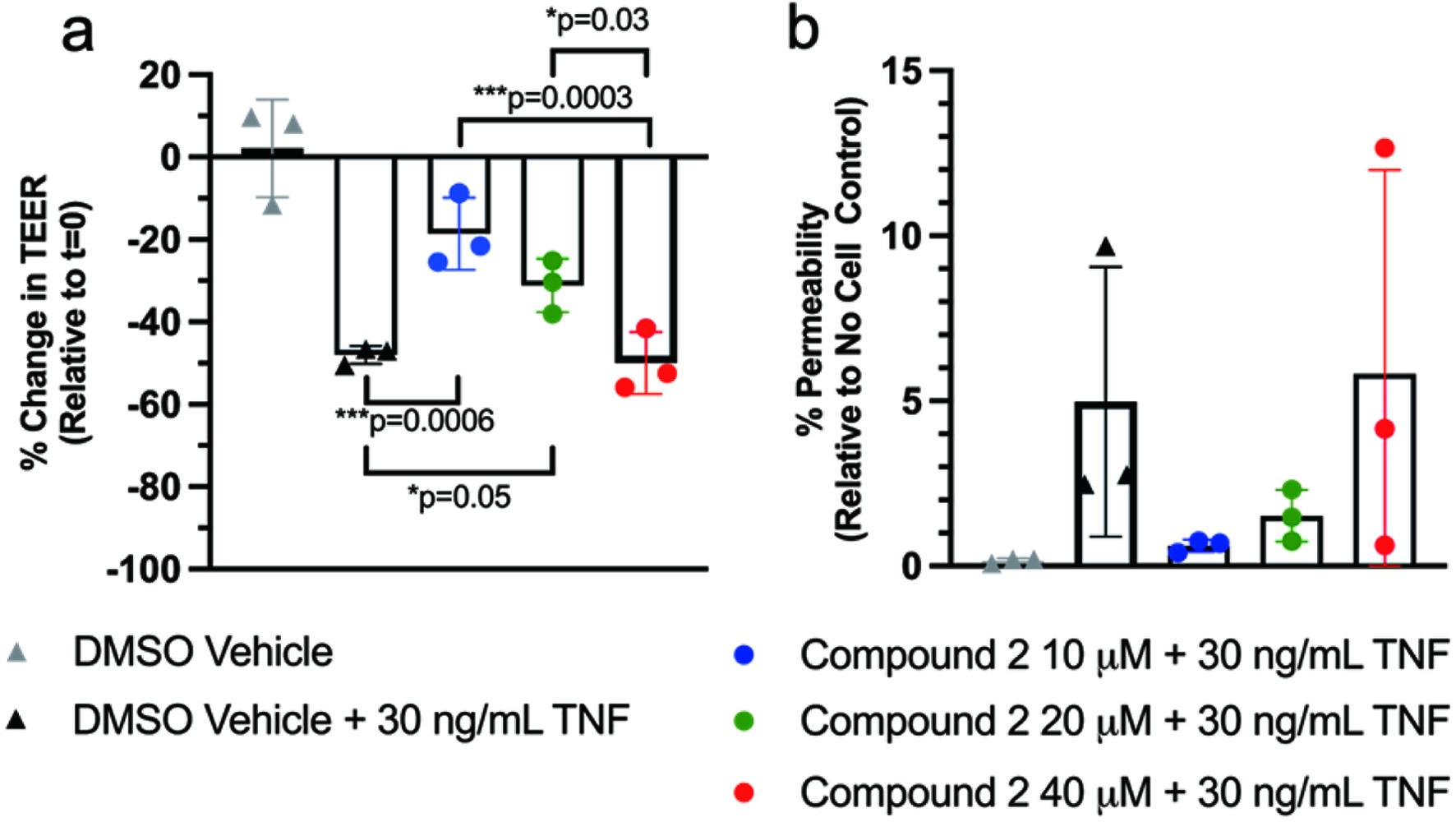 Click for large image | Figure 3. Administration of NFT protected against (a) decreased TEER and (b) increased permeability with TNF-induced inflammation. Compound 2 refers to NFT. |
When administered in combination, NCT and NFT exhibited the greatest beneficial effect at 20 μM with respect to reversing TEER changes due to TNF (Figure 4a). The lowest doses (10 μM, 20 μM) had the greatest benefit on % permeability, while the highest dose (40 μM) did not yield a substantive change in % permeability compared to control (Figure 4b).
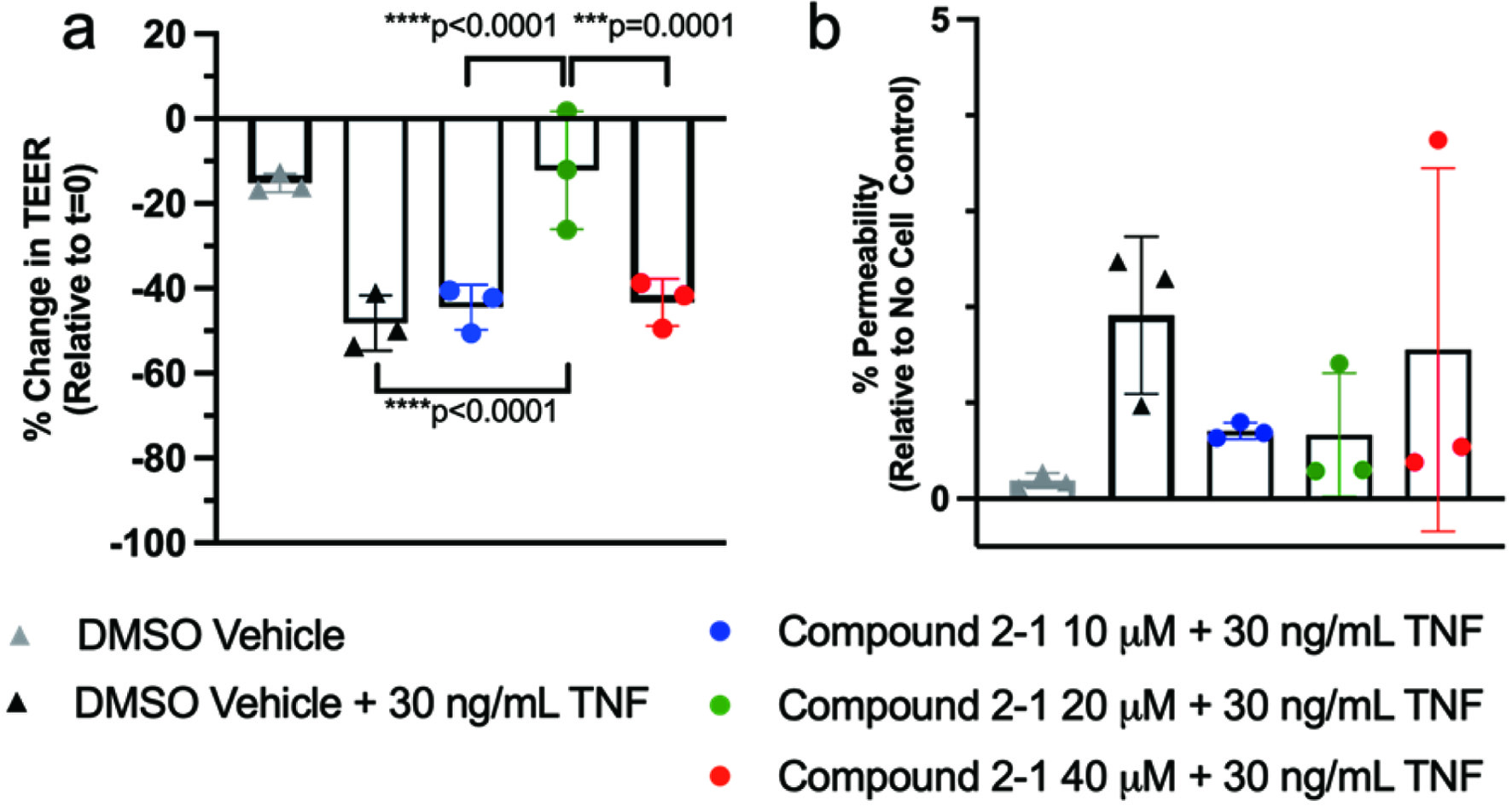 Click for large image | Figure 4. Administration of NCT:NFT (a) modulated TEER and (b) reduces permeability with TNF-induced inflammation. Compound 2-1 refers to a combination of NCT and NFT. |
| 4. Conclusion | ▴Top |
Current data on NCT and NFT indicate that its role as an HNF4α agonist can affect many organs and organ systems. The data presented in this contribution show that NCT, and, to a lesser extent, NFT, positively affect gut barrier function in a state of elevated inflammation. This is an example of leveraging cutting-edge technology (artificial intelligence) to identify key compounds, among thousands of candidate bioactive compounds, that have a targeted effect at the molecular level.
The current body of literature demonstrates NCT and NFT’s role in liver health and lipid flux within the liver, as well as gut health (Veeriah et al., 2022). Previous work has demonstrated a role for NCT and NFT in gut barrier function due to its HNF4α-agonist properties. Herein, we confirm that NCT and NFT improve markers of gut barrier function during a state of elevated inflammation in vitro. These two bioactive compounds have the potential to exert profound effects on human health. However, as with most bioactive compounds, there are few recommendations for daily intake. As many countries look for solutions to address rising health care costs and overall health and well-being, bioactive compounds may hold the key to reducing reliance on conventional treatment approaches and prioritizing prevention through plant-derived nutritional interventions. The Academy of Nutrition and Dietetics’ recommendation for daily flavan-3-ol intake is the first of its kind from an authoritative health and nutrition organization. Between the growing body of evidence that supports the critical role of bioactive compounds in human health and the first of its kind recommendation for daily intake, we are at a turning point for more research and recommendations for bioactive compounds in the daily diet.
Several pre-clinical experiments, including the data presented in this contribution, have demonstrated a potential for NCT to exert a HNF4α-mediated benefit on intestinal function. Additionally, HNF4α has a well-documented effect on epithelial cell transcription activity and the related transcriptome (Ntunzwenimana et al., 2021). In patients with ulcerative colitis, HNF4α is significantly downregulated compared to controls, suggesting a role in the disease state (Vancamelbeke et al., 2017). Thus, the case for HNF4α’s role in gut inflammation and overall gut health is evident.
NCT and NFT have the potential to mediate gut health and gut barrier integrity via HNF4α. Additional work is underway to validate this hypothesis. Additional clinical research is warranted to demonstrate the aforementioned benefits in humans. The evidence is mounting for an evolved nutrition framework that places bioactive compounds alongside traditional nutrients like vitamins and minerals. The inclusion of bioactive compounds in dietary recommendations can inform a new, national standard for nutrition and dietary guidelines and enable a new food-as-medicine paradigm for preventative health.
Acknowledgments
The authors would like to thank Robert Fultz, PhD, Biocuration Scientist, Brightseed for biostats support and Altis Biosystems for their services. This study was funded by Brightseed. Manuscript preparation supported by Maria Stewart, PhD (Growing Brilliance LLC).
| References | ▴Top |