| Journal of Food Bioactives, ISSN 2637-8752 print, 2637-8779 online |
| Journal website www.isnff-jfb.com |
Review
Volume 18, June 2022, pages 1-42
SARS-CoV-2-induced host metabolic reprogram (HMR): nutritional interventions for global management of COVID-19 and post-acute sequelae of COVID-19 (PASC)
A. Satyanarayan Naidua, *, Fereidoon Shahidib, Chin-Kun Wangc, Kenji Satod, Aman Wirakartakusumahe, Ogugua C. Aworhf, Roger A. Clemensg
aN-terminus Research Laboratory, 232659 Via del Rio, Yorba Linda, CA 92887, USA
bDepartment of Biochemistry, Memorial University of Newfoundland, St John’s, NL, A1C 5S7, Canada
cSchool of Nutrition, Chung Shan Medical University, 110, Section 1, Jianguo North Road, Taichung 40201, Taiwan
dDivision of Applied Biosciences, Graduate School of Agriculture, Kyoto University, Kitashirakawa Oiwake-cho, Sakyo-ku, Kyoto 606-8502, Japan
eDepartment of Food Science and Technology, Bogor Agricultural University (IPB), Indonesia
fDepartment of Food Technology, University of lbadan, Nigeria
gUniversity of Southern California, School of Pharmacy, D. K. Kim International Center for Regulatory Science, 1540 Alcazar St., CHP 140, Los Angeles, California 90089, USA
*Corresponding author: A. Satyanarayan Naidu, N-terminus Research Laboratory, 232659 Via del Rio, Yorba Linda, CA 92887, USA. E-mail: asnaidu@nterminus.com
DOI: 10.31665/JFB.2022.18306
Received: June 2, 2022
Revised received & accepted: June 30, 2022
| Abstract | ▴Top |
‘Severe acute respiratory syndrome coronavirus 2’ (SARS-CoV-2) is a highly transmissible viral pathogen responsible for the ongoing ‘coronavirus disease 2019’ (COVID-19) pandemic. The current re-purposed antiviral interventions against SARS-CoV-2 are classified into two major groups: Group-1 represents the family of drugs, mainly the vaccines that directly target the virus, and Group-2 includes a specific class of inhibitors that interfere with the host-cell machinery, which is critical for viral infection and replication. Global efforts to control COVID-19 pandemic with vaccines and repurposed therapeutics represent only a phased victory. The emergence of several SARS-CoV-2 variants of concern (VOCs) has compromised several vaccinations and pharma-therapeutic protocols, which highlights the dire necessity for specific antiviral interventions that target highly conserved domains, which are less likely to mutate in the SARS-CoV-2 genome. Several bioactive phytochemicals that block viral enzymes such as nsp5/main proteinase (Mpro) and RNA-dependent nsp7/nsp8/nsp12 RNA-dependent RNA-polymerase (RdRp) complex, are extensively investigated in this direction. The SARS-CoV-2 infection triggers a complex human host-pathogen interaction(s) resulting in ‘host metabolic reprogramming’ (HMR), iron (Fe)-redox dysregulation (FeRD), and altered mitochondrial function that cumulatively disrupt several metabolic pathways involved in cellular energy and antioxidant enzyme function; thereby, compromise the innate host defense. The circulatory/RAAS axis contributes to FeRD and any alteration or imbalance in the Fe-redox homeostasis (Fe-R-H) may lead to ‘new onset’ metabolic disorders (i.e., diabetes). Such inherent body damage and its long-term health consequences in post-acute sequelae of COVID-19 (PASC) require effective nutritional intervention strategies, particularly at the interface of organ system functions and immune system dynamics. The long-term sequelae of PASC indicate an accelerated rate of immune exhaustion in COVID-19 patients, due to prolonged antigen stimulation (also due to vaccine exposure). Abnormal immune metabolism may also cause systemic perturbations (i.e., FeRD), ROS/RNS production, oxidative and nitrosative stress, which could trigger multi-organ disorders ranging from mild symptoms to an incapacitating state and reduced quality of life that could last for weeks or longer following recovery from COVID-19. The five most long-term clinical manifestations of PASC include fatigue, headache, attention disorder, hair loss, and dyspnea. This narrative review elucidates the intricate impairments and sequelae associated with eight major physiological systems in COVID-19 survivors (i.e., pulmonary, neuro-cognitive, cardiovascular, renal, gastrointestinal/hepato-biliary, endocrinal, skeleton-muscular, and reproductive)—triggered by the FeRD, amplified by the HMR, altered mitochondrial function and ACE2/RAAS axis. We have attempted to explain the ongoing epidemic of the residual, non-viral host metabolic disorders and complications in COVID-19 survivors and the supportive role of specific host system-targeted nutritional interventions such as natural plant-based anti-inflammatories, immune-modulators, antioxidants, and macro-/micronutrient metabolic optimizers to manage PASC, the newly emerged post-COVID metabolic syndrome.
Keywords: COVID-19; Post-Acute Sequelae of COVID-19 (PASC); Host Metabolic Reprogramming (HMR); Iron (Fe)-Redox Dysregulation (FeRD); Food Bioactives; Nutritional Interventions
| 1. Introduction | ▴Top |
‘Severe acute respiratory syndrome coronavirus 2’ (SARS-CoV-2) is a highly transmissible viral pathogen responsible for the ongoing ‘coronavirus disease 2019’ (COVID-19) pandemic. The COVID-19 pandemic continues to affect millions of people worldwide. The current global statistics include 540 million cases, about 6.3 million deaths (WHO, 2022). The SARS-CoV-2 is a single-stranded positive-sense RNA virus with high mutation rate (up to a million-fold higher than their hosts) and such extreme genetic rearrangement correlates with enhanced adaptability and virulence, a trait that works for the viral advantage (Duffy, 2018). Its replication machinery is highly error-prone without correction systems; therefore, coronaviruses (CoVs) are prone to several genetic alterations during an infectious life cycle (Perales and Domingo, 2016). Accordingly, SARS-CoV-2 could rapidly evolve, pose high risk of transmission, and frequently develop drug resistance as well as evade vaccine-induced immunity (Duffy, 2018; Pruijssers and Denison, 2019). Such genomic advantage makes the SARS-CoV-2 pathogen a major challenge to develop effective antiviral strategies and control COVID-19.
Considering that vaccination is an effective strategy to control COVID-19 pandemic, a massive public health campaign in history, has administered more than 11.9 billion doses across 185 countries with a rate of about 19.3 million vaccinations per day (WHO, 2022). However, the ability of SARS-CoV-2 to evade vaccine-induced immunity—has become a global concern; since a large population has been vaccinated and the pressure of herd immunity could force genomic adaptation to evolve novel viral variants as ‘escape’ mutants. Such genetic drift in tandem with the evasion of immune recognition, has contributed to the emergence of several SARS-CoV-2 variants of concern (VOCs) (Koyama et al., 2020; Naidu et al., 2022a). Accordingly, the recent spread of Omicron variant (B.1.1.529) compromised several vaccination and public health safety protocols (Minka and Minka, 2022). These VOCs show increased transmissibility and/or immune evasion, traits that are linked to mutations in the viral spike (S) protein (Harvey et al., 2021; Tao et al., 2021). Such high number of mutations in the S-protein domain raises concern that this CoV pathogen could evade antibodies elicited by natural infection or vaccination and the therapeutic monoclonal antibodies may also become less effective (Hoffmann et al., 2022). There is a desperate need for an effective COVID-19 vaccine to control rapid transmission of the viral pathogen, and to contain the emergence of potential SARS-CoV-2 variants. Unfortunately, there is no effective ‘multivalent vaccine’ yet that could provide immune protection against multiple SARS-CoV-2 variants (Naidu et al., 2022a).
The genomic size of an organism is inversely related to the error rate during replication, also in case of SARS-CoV-2 (RNA length ∼30 kb), which translates to one nucleotide substitution for every 2-3 genomes synthesized (Swanstrom and Schinazi, 2022). In general, most mutations are deleterious; however, a subset of mutations may potentially give rise to a viral phenotype (Holmes, 2011), such as the VOC Omicron (B 1.1.529). A typical SARS-CoV-2 infected individual is estimated to produce about 1-100 billion virions during peak phase of CoVID-19 (Sender et al., 2021). Taken together, its high viral replication rate in tandem with inherent genetic mutability, its compact genomic size with lack of metabolic machinery makes SARS-CoV-2 an extremely challenging druggable antiviral target. Current global health crisis highlights the dire necessity for specific antiviral intervention strategies that target highly conserved domains, which are less likely to mutate in the SARS-CoV-2 genome (Krumm et al., 2021).
| 2. Viral pathogen-targeted COVID-19 interventions | ▴Top |
The current vaccines and re-purposed antiviral interventions against SARS-CoV-2 could be classified into two major groups: Group-1 are family of drugs, mainly the vaccines that directly target the virus, and Group-2 includes the type of inhibitors that interfere with host-cell machinery critical for viral infection and replication.
2.1. Group-1: COVID-19 vaccines
Four COVID-19 vaccines, two mRNA-based: BNT162b2 (Pfizer-BioNTech) and mRNA-1273 (Moderna), and two adenoviral vector-based: Ad26.COV2.S (Janssen/Johnson & Johnson) and ChAdOx1 nCoV-19 (Oxford/AstraZeneca) have been approved or granted Emergency Use Authorization (EUA) for COVID-19 control in many nations worldwide. The ChAdOx1 nCoV-19 vaccine has not yet received a EUA or approval from the US-FDA (US-FDA/EUA, 2022).
2.1.1. COVID-19 vaccines and the ‘original antigenic sin’
Based on meta-transcriptome sequencing of the bronchoalveolar lavage fluid from COVID-19 patients, the viral pathogen seems to evolve in vivo after infection, a characteristic that may determine its virulence, infectivity, and transmissibility (Roncati and Palmieri, 2020). Therefore, this unprecedented race to develop COVID-19 vaccine should follow caution that the viral antigen candidates are safe and not detrimental to host immune responses. Immune enhancement, also known as ‘immune backfiring’, could manifest in multiple ways such as antibody-dependent enhancement (ADE), a process in which a virus could leverage antibodies to aid infection; or cell-based enhancement, a category that includes allergic inflammation caused by Th2 immunopathology (Peeples, 2020). The anecdotal reports of COVID-19 reinfections may suggest the relevance of ADE, where the viral antibodies (from immunization or an initial natural infection) might have enhanced the viral entry into host cells (Lan et al., 2020).
2.1.2. Covid-19 vaccine and autoimmune sequelae
During autoinflammatory and autoimmune syndromes, viruses may activate an aberrant innate and acquired immune response, with increased synthesis of cytokines, mainly TNF-α, IL-6 and IL-1β, IL-17, IL-18, in genetically predisposed individuals (Caso et al., 2020). Such immune hyperactivation and excess cytokine release is evident in COVID-19 patients with multi-organ failure and fatal outcomes (Rodríguez and Brodin, 2020a). Molecular mimicry between SARS-CoV-2 antigens and the human proteome could also play an important role is this response (Vojdani and Kharrazian, 2020). Thus, after an infection or vaccination, the host immune responses elicited by SARS-CoV-2 antigenic epitopes may cross-react with human proteins that share peptide sequences and trigger severe autoimmune sequelae. From a clinical context, SARS-CoV-2 shares 6 minimal immune determinants with the Kawasaki antigen inositol-trisphosphate 3-kinase C that could predispose likely cross-reactions and consequent autoimmune Kawasaki disease in COVID-19 patients (Ehrenfeld et al., 2020). Moreover, the SARS-CoV-2 spike protein has been shown to share 13 out of 24 pentapeptides homologous to the human lung surfactant proteins (Kanduc and Shoenfeld, 2020). Therefore, identification of human tissue cross-reactive epitopes in COVID-19 vaccine is critical to avoid any possible autoimmune sequelae. Only peptide sequences unique to SARS-CoV-2 could represent the basis for safe and specific vaccination protocols.
COVID-19 vaccine-induced endocrine disorders include vaccine-associated thrombosis and thrombocytopenia (TTS) with adrenal hemorrhage, derangements in glycemic control including new onset type 2 diabetes mellitus, and subacute thyroiditis (Mirza et al., 2022). TTS is a serious but rare adverse event associated with exposure to adenovirus vector vaccines Ad26.COV2.S and ChAdOx1 nCoV-19, which has raised immunization safety concerns (Long et al., 2021; See et al., 2021). Symptoms of such novel vaccine-induced clinical syndrome include severe headache, blurred vision, seizure, severe and persistent abdominal pain, painful swelling of the lower leg, and chest pain or dyspnea. The TTS mimics autoimmune heparin-induced thrombocytopenia (HIT) mediated by platelet-activating antibodies against platelet factor 4 (Lai et al., 2021). It is also known as vaccine-induced prothrombotic immune thrombocytopenia (VIPIT) in some European nations and Canada (Aleem and Nadeem, 2022). A systematic review of 160 cases from 16 countries, revealed that the TTS onset occurs at a median of 9 (4) days after vaccination with a high mortality rate of 36.2% (Waqar et al., 2021). Venous thrombosis (61%) in TTS is prevalent and about 66.3% affected could develop cerebral venous sinus thrombosis (CVST), predominantly among the female patients (aged <55 years). By April 12, 2021, ∼7 million doses of Ad26.COV2.S vaccine were given in the US, with 6 cases of CVST with thrombocytopenia were reported, which called for a temporary national pause in vaccination with Ad26.COV2.S on April 13, 2021. The initial 12 US cases of CVST with thrombocytopenia after vaccination resulted in severe outcomes (See et al., 2021). Newly emerged TTS is a major concern for global implementation of mass COVID-19 vaccination campaigns and requires stringent caution with ongoing COVID-19 vaccine development protocols.
2.2. Group-2: COVID-19 antiviral drugs
Specific novel anti-SARS-CoV-2 medications are anticipated to play a major role in protecting unvaccinated or immunocompromised individuals, as well as at periods when vaccinations fail to protect against circulating SARS-CoV-2 variant (Chavda et al., 2021). The antiviral strategies for COVID-19 control fall under four categories: i) inhibitors of viral entry, bioactives that block human cell surface receptors such as ACE2 (TMPRSS2), neuropilin-1, and heparan sulfate that SARS-CoV-2 uses for cellular invasion; ii) inhibitors of viral proteases (i.e., Mpro, RdRp), enzymes that hydrolyze long viral polypeptides to generate functional proteins; iii) inhibitors of viral replication, transcription, and translation, i.e., nucleoside analogs that mimic RNA bases that a virus could potentially incorporate into copies of it’s genome; and iv) inhibitors of viral assembly and release, i.e., bioactives that target host cellular processes that are hijacked for transport and assembly of viral particles.
2.2.1. Repurposed drug therapeutics
In the U.S., the Coronavirus Treatment Acceleration Program (CTAP) initiated by the FDA, is an emergency plan to scrutinize and introduce new effective therapeutics to COVID-19 patients, supported by extensive safety and efficacy evaluation (US-FDA/CTAP, 2022). As of March 19, 2022, the CTAP review included (vaccines excluded) over 690 drug development programs (in planning stages), over 470 trials (under review), about 15 COVID-19 treatments (authorized for emergency use) and only 1 antiviral drug (remdesivir) approved. Currently, the diversity of COVID-19 interventional strategies under CTAP investigation include: 50+ antiviral treatments, 60+ cells/gene therapies, 120+ immunomodulators, 60+ neutralizing antibodies, 110+ other interventions and 40+ combination therapeutics (from other drug categories) (US-FDA, 2022). As of March 2022, over 7,710 ongoing clinical trials were registered on ClinicalTrials.gov, which include 1,252 vaccine-related, 1,120 drug intervention and 155 dietary supplement studies (NIH/US-NLM, 2022).
2.2.2. Current status
Despite robust efficacy in vitro data against the SARS-CoV-2 pathogen and previous clinical data from other human CoVs such as SARS and MERS, the repurposed drugs have failed to demonstrate beneficial effects against COVID-19 in human clinical studies (Indari et al., 2021; Martinez et al., 2021; Basu et al., 2022).
| 3. Human host-targeted COVID-19 management strategies | ▴Top |
The virulence potential of SARS-CoV-2 to invade a wide range of cells and tissues beyond the respiratory system, is manifested into a broad range of clinical syndromes (i.e., FeRD, ARDS, SIRS, AI, etc.), with varying degrees of severity ranging from asymptomatic, mild, moderate, to severe fatal multi-organ dysfunction syndrome (MODS). The possible risk of a long-term damage to certain affected host organ/systems or the elevated risk of disorders in later life could significantly worsen the burden on global healthcare. Considering the broad diversity of clinical symptoms, populations, and underlying comorbidities there is dire necessity to develop human host-targeted clinical management strategies to combat COVID-19 pandemic.
3.1. Host metabolic reprogramming (HMR) in COVID-19
Viruses hijack the host cellular metabolic machinery to extract adequate energy and carbon skeletons required for their entry and further molecular constructions of viral progeny inside a host cell. The SARS-CoV-2 infection could activate a complex human host-pathogen interactions leading to host metabolic reprogramming (HMR). The HMR could alter mitochondrial function with significant disruption of glycolysis/tricarboxylic acid (TCA) cycle affecting several metabolic pathways of amino acid, fatty acid, nucleotide, and antioxidant synthesis (Moolamalla et al., 2021; Shen and Wang, 2021). The impact of HMR on COVID-19 pathobiology is reflected during the hyper-inflammatory response (‘cytokine storm’) while compromising the innate host defense. This could sequentially trigger an array of clinical manifestations, either an asymptomatic condition or progressive onset of mild, moderate to severe phases of COVID-19 with life-threatening acute respiratory distress syndrome (ARDS), vascular dysfunction, multiple-organ failure, and death. Therefore, nutritional restoration of HMR could provide a potential strategy to combat COVID-19 and its post sequelae.
3.2. Tri-phasic symptomatic progression of COVID-19
The symptomatic progression of COVID-19 requires that a genetically competent (virulent) SARS-CoV-2—i) infects and invades a susceptible host via specific cell surface receptors, ii) induces HMR to ensure ready access to an active host cellular machinery for an uninterrupted viral propagation, iii) inactivates innate host defense to evade viral elimination, and iv) exits the infected host cell and repeats the viral propagation cycle for exponential growth and transmission (Sicari et al., 2020). In accordance with its virulence spectrum and host susceptibility pattern, the symptomatic outcomes of COVID-19 manifest in a tri-phasic manner as iron (Fe)-redox disruptive hematological syndromes (Naidu et al., 2022a) (Figure 1).
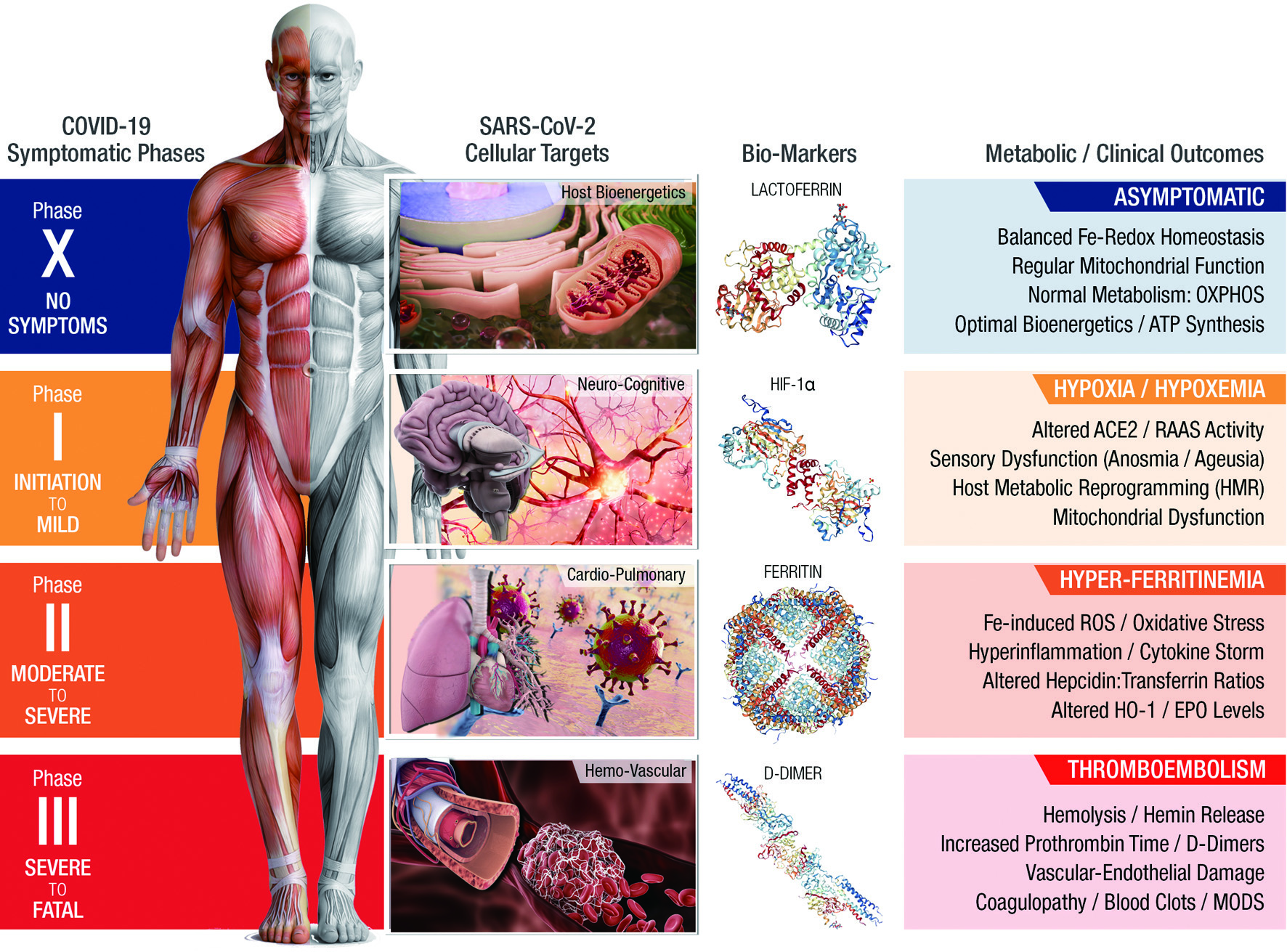 Click for large image | Figure 1. COVID-19: an iron (Fe)-Redox Dysregulation (FeRD) Syndrome. A SARS-CoV-2-induced host metabolic reprogramming (HMR). |
3.2.1. Phase-I/hypoxia/hypoxemia (an acute depletion of oxygen (O2) transport in the blood)
During the initial encounter, the SARS-CoV-2 anchors to the human angiotensin-converting enzyme 2 (ACE2) receptors on alveolar epithelia, alters the renin-angiotensin-aldosterone system (RAAS), subsequently lowers both blood pressure and lung function of the infected host (Ni et al., 2020). Reduced O2 transport that ensues from low hemoglobin (Hb) levels in the hypoxic blood circulation (‘hypoxemia’), alters the mitochondrial function by ‘switching off’ the oxidative phosphorylation (OXPHOS)/TCA cycle in favor of anaerobic glycolysis (Ferraro et al., 2021). Therefore, beyond the classical pulmonary immune-hyperinflammation and ARDS, COVID-19 manifests also as a hypoxia-induced hematological syndrome with ‘iron-related’ HMR (Debuc and Smadja, 2021).
3.2.2. Phase-II/hyperferritinemia (an excess presence of iron storage protein, ferritin, in the blood)
In its subsequent infectious phase, SARS-CoV-2 induces hyper-release of proinflammatory cytokines to stimulate synthesis of both ferritin and hepcidin, the ultimate mediators of iron dysregulation (Edeas et al., 2020). This pathological condition is reflected by high iron content in reticuloendothelial cells with elevated serum ferritin levels. Excess iron load further generates reactive oxygen species (ROS) through Haber-Weiss reaction, which leads to oxidative stress, mitochondrial dysfunction and ferroptosis (Singh et al., 2020c). Taken together, hyperferritinemia, cellular imbalance in iron metabolism plays a critical role in the pathogenesis of COVID-19 (Muhoberac, 2020).
3.2.3. Phase-III/Thromboembolism (formation of blood clots with severe obstruction of veins, arteries, and circulation)
During this severe stage of COVID-19, hematological parameters such as anemia of inflammation (AI), low counts of peripheral blood lymphocytes/eosinophils with increased polymorphonuclear-to-lymphocyte ratios are prominent risk factors (Sun et al., 2020; Bergamaschi et al., 2021). Altered iron metabolism, iron-restricted erythropoiesis due to hyper-inflammation are predisposing factors for AI (Wessling-Resnick, 2018; Weiss et al., 2017). The hemolysis-derived heme could initiate oxidative and inflammatory stress that may lead to microvascular thrombosis, organ ischemia and multi-organ failure in severe COVID-19 cases (Wagener et al., 2020; Varga et al., 2020).
3.3. Host Iron (Fe)-redox dysregulation (FeRD) in COVID-19
The significant role of iron metabolism in HMR and altered mitochondrial bioenergetics is evident in the pathobiology of COVID-19 (Terpos et al., 2020; Naidu et al., 2021c). Throughout the tri-phasic clinical progression of COVID-19, the SARS-CoV-2 pathogen categorically targets the host hematopoietic system and alters the host ‘iron (Fe)-redox homoeostasis (Fe-R-H)’ (Naidu et al., 2022a). The Fe-redox dysregulation (FeRD) could also trigger several clinical manifestations in COVID-19 patients including: i) decrease the functional hemoglobin (Hb), ii) increase the cellular iron overload, iii) release free toxic heme into the circulation, iv) manifest hypoxemia and systemic hypoxia, v) reduce nitric oxide (NO•) synthesis, vi) activate coagulation pathway(s), vii) trigger ferroptosis with oxidative stress and lipoperoxidation, and viii) induce mitochondrial degeneration (Cavezzi et al., 2020). Therefore, regulation and maintenance of systemic Fe-R-H is critical for the clinical management of COVID-19.
Interestingly, FeRD and its associated physiological disorders or disease states continue for extended periods (for weeks or even months) in COVID-19 patients discharged as RT-PCR (SARS-CoV-2) negative survivors (Taribagil et al., 2021). These observations further emphasize the need to identify intricate pathophysiological mechanisms underlying FeRD condition in COVID-19. Based on the consequential clinical manifestation of SARS-CoV-2 infection (i.e., COVID-19) and the ‘post-acute sequelae of COVID-19’ (PASC), this disease should be considered as an Iron (Fe)-Redox Dysregulation (FeRD) Syndrome.
A robust correlation between COVID-19 and the host Fe-R-H dysregulation is also observed among specific population groups with hemoglobin (Hb) anomalies (Rapozzi et al., 2021; Naidu et al., 2022a). Global data indicate higher COVID-19 case fatality rates (CFR) among men than women, a ratio >1.0, ranging up to 3.5 in some cases (Global Health, 2022), a gender trait attributed to low Hb levels in females compared to males (Conti and Younes, 2020; Jin et al., 2020). Also, most newborns from COVID-19 positive mothers remain uninfected with the virus (Naidu et al., 2022c), which could be related to the absence (and delayed synthesis) of Hb β-chain in neonatal erythropoiesis (Sankaran and Orkin, 2013). Interestingly, the low incidence of COVID-19 cases reported from certain Mediterranean territories is noteworthy, since this geographical region is known for high prevalence of β-thalassemia (blood condition linked to abnormalities in the β-chains of Hb) (Drouin, 2020; Motta et al., 2020).
| 4. Nutritional strategies to reverse FeRD condition in COVID-19 | ▴Top |
Severe imbalance in iron metabolism (Fe-R-H dysregulation) is prominent in every symptomatic (mild, moderate to severe) clinical phase of COVID-19. Functional iron deficiency and Fe-R-H are reported in 80% of COVID-19 patients, and the advanced anemia of inflammation (AI) is associated with significantly longer hospital stay and increased CFR. Notably, the recovery of COVID-19 patient results in resolution of anemia and normalization of dysregulated Fe-R-H (Lanser et al., 2021). The Fe-R-H dysregulation with elevated ferritin/transferrin ratio predicts insufficient pulmonary oxygenation with the need for ICU admission and mechanical ventilation for COVD-19 patients (Bellmann-Weiler et al., 2020). Therefore, Fe-R-H restoration is a host biomarker-driven potential combat strategy for an effective clinical and post-recovery management of COVID-19 (Naidu et al., 2022a). The following section describes nutritional strategies, using Fe-R-H regulators, ferroptosis inhibitors, anticoagulants, and iron chelators to reverse host Fe-R-H dysregulation in COVID-19 patients (Figure 2).
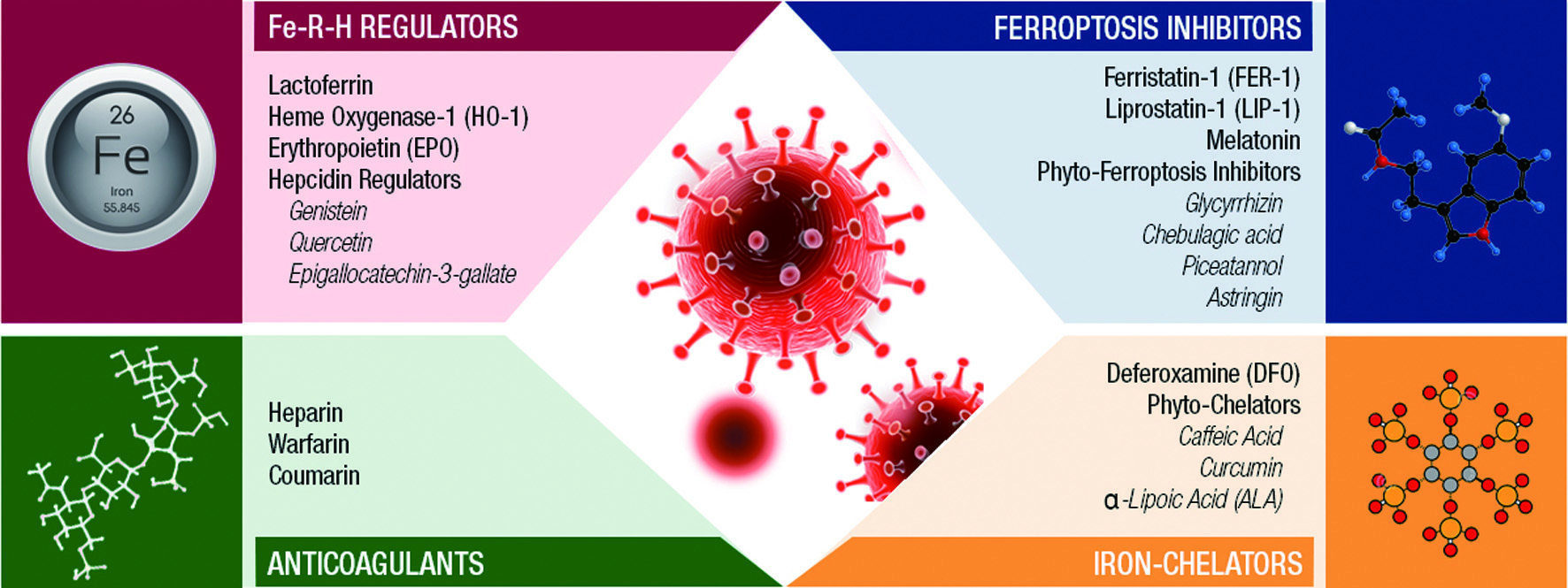 Click for large image | Figure 2. Nutritional strategies to reverse host Fe-R-H dysregulation in COVID-19. |
4.1. Fe-R-H regulators for nutritional management of COVID-19
The Fe-R-H regulators, such as lactoferrin (LF), hemeoxygenase-1 (HO-1), erythropoietin (EPO), and hepcidin modulators are innate bioactive molecules involved in iron metabolism, detoxification of free iron-induced ROS, modulation of antioxidant responses and serve as the first barriers against SARS-CoV-2 infection (Naidu et al., 2022a). These Fe-R-H regulators could play a vital role in alleviating cellular oxidative stress and inflammation, particularly during ‘cytokine storm’ in COVID-19 pathobiology.
Lactoferrin, the iron-binding protein found in milk and several exocrine secretions, could also interact with anionic compounds i.e., heparan sulfate proteoglycan (HSPG), ACE2 etc., translocate into the nucleus, modulate immune as well as inflammatory responses, and regulate the physiological Fe-R-H status (Naidu et al., 2022a). Accordingly, LF is considered a potential natural intervention in the clinical management of COVID-19 (Chang et al., 2020; Campione et al., 2021; Naidu et al., 2022b). Based on iron sequestration and Fe-R-H restoration effects in vivo, LF could potentially benefit COVID-19 patients to relieve oxidative stress and hyper-inflammation (Naidu et al., 2021c). Several LF-based intervention technologies are undergoing extensive clinical trials for COVID-19 control.
Hemeoxygenase-1 (HO-1) regulates the Fe-R-H and provides cytoprotective function via endogenous mechanisms to sustain body’s antioxidant response against oxidative stress (Consoli et al., 2021). The protective role of HO-1 against SARS-CoV-2 is probably an emergency inducible defense mechanism to ameliorate oxidative stress from heme-released oxidants in severe cases of COVID-19. Several natural phytochemicals are known to upregulate HO-1; thus, may provide protection from thrombotic events and vascular inflammation during COVID-19 (Naidu et al., 2022a). Nimbolide, a limonoid tetranortriterpenoid from neem plant (Azadirachta indica), resveratrol (3,4′,5-trihydroxy stilbene) and curcuminoids (with α, β-unsaturated carbonyl groups) are potential inducers of HO-1 expression through Nrf2/antioxidant-responsive element (ARE) pathway (Chen et al., 2005; Jeong et al., 2009; Mahapatra et al., 2012). The phytophenolic compound quercetin, could induce HO-1 expression via mitogen-activated protein kinase (MAPK)/Nrf2 pathway (Yao et al., 2007).
Erythropoietin (EPO) could protect pulmonary vascular beds and counteract hypoxic pulmonary vasoconstriction (Nairz et al., 2012). Neurological manifestations are prominent among COVID-19 patients, Also, EPO could relieve acute and chronic-progressive downstream sequelae of central and peripheral nervous systems, which are now becoming common neurological manifestations among COVID-19 patients (Naidu et al., 2021c; Collantes et al., 2021). A combination therapy of EPO with anti-coagulants or anti-thrombotic agents (i.e., heparin) could circumvent complications in hospitalized COVID-19 patients (Nairz et al., 2012).
Hepcidin is a promising intervention target for COVID-19 control with iron overload syndromes. (Blanchette et al., 2016; Banchini et al., 2020). Several dietary phytoestrogens could up-regulate hepcidin expression, control systemic iron levels, prevent iron-induced toxicity and provide protection against several oxidative stress-induced pathological disorders (Bayele et al., 2015). Genistein, the isoflavone-related estrogen could induce hepcidin transcription by both bone morphogenetic protein 6 (BMP6) and signal transducer and activator of transcription 3 (STAT3) signaling (Zhen et al., 2013). Quercetin, a strong post-prandial hepcidin inducer, has been shown to reduce iron overload (Kaltwasser et al., 1998). Epigallocatechin-3-gallate (EGCG) may reduce iron toxicity by chelation or blocking the iron release/efflux from cells (Perron and Brumaghim, 2009). As potential inducer of hepcidin expression, phytoestrogens are promising adjunctive supplements to reduce iron overload and prevent any sequelae of iron-induced toxicities such as hyperferritinemia, coagulopathies and/or thromboembolism, the prominent clinical manifestations in COVID-19 patients (Sonnweber et al., 2020; Edeas et al., 2020; Habib et al., 2021).
4.2. Ferroptosis inhibitors for nutritional management of COVID-19
Ferroptosis is an iron-catalyzed, non-apoptotic form of regulated necrosis that causes oxidative damage of cellular lipid membranes leading to severe mitochondrial dysfunction. The ferroptosis-mediated FeRD could cause suppression of erythropoiesis and anemia, which is a prominent feature in severe cases of COVID-19. During ferroptosis, the accumulation of oxidized phospholipids in myocardial and renal tissues cause ischemic-reperfusion injury, a detrimental factor for cardiac damage and MODS in COVID-19 (Jacobs et al., 2020). Ferroptosis is linked to several neurological disturbances including cognitive impairment, ageusia and anosmia in COVID-19 (Vaira et al., 2020b). Ferroptosis could also be involved in the development of acute lung injury/acute respiratory distress syndrome (ALI/ARDS), a major contributor for high morbidity and mortality in COVID-19 (Liu et al., 2022). Therefore, ferroptosis inhibitors could provide a potential intervention strategy to alleviate thromboembolism and improve prognosis in COVID-19 patients (Yang and Lai, 2020b; Naidu et al., 2022a).
The pharmaceutical drugs ferrostatin-1 (Fer-1) and liproxstatin-1 (Lip-1) are potent lipophilic free-radical scavengers that could prevent lipid peroxidation and protect from ferroptosis. Melatonin inhibits platelet activation and ferroptosis through activation of Nrf2 and HO-1 signaling pathways (Ma et al., 2020). This chronobiotic hormone is considered a potential intervention to treat hemolytic, thrombotic, and thrombocytopenic conditions, the widespread clinical manifestations among COVID-19 patients (Naveenkumar et al., 2019; Naidu et al., 2022a).
Several phytochemicals are potential ferroptosis inhibitors. The phytoflavone quercetin could upregulate cellular GSH and inhibit ferroptosis of renal proximal tubular epithelia by reducing malondialdehyde (MDA) and oxidative damage of lipid membranes (Wang et al., 2020b). Similarly, curcumin could inhibit renal tubular epithelial ferroptosis from myoglobin-mediated inflammation and oxidative stress through activation of cytoprotective enzyme HO-1 (Guerrero-Hue et al., 2019). Glycyrrhizin from licorice (Glycyrrhiza glabra) could provide anti-ferroptotic liver protection through up-regulation of Nrf2, and HO-1 and down-regulation of lactate dehydrogenase (LDH), MDA, and free iron (Wang et al., 2019). Phyto-tannins chebulagic and chebulinic acids are natural iron-chelators that inhibit ferroptosis through free radical scavenging and other regular antioxidant pathways (Yang et al., 2021a). Dietary phytophenols such as piceatannol and astringin strongly inhibit ferroptosis via preferential transfer of hydrogen (H+) atoms as conventional antioxidants. (Chen et al., 2021a).
4.3. Anticoagulants for COVID-19 management
The severe phase of COVID-19 is a highly prothrombotic disease state resulting from hyper-inflammation, endothelial dysfunction, platelet and complement activation, derangement of RAAS system, and hypoxemia. The ARDS and multi-organ failure in COVD-19 have been attributed to markers of coagulopathy such as prothrombin (PT) prolongation, elevated fibrin degradation products, reduced platelet count, and significantly elevated D-dimers (Giannis et al., 2020; Tang et al., 2020). Therefore, thromboprophylaxis with anticoagulant therapy has been widely practiced as a COVID-19 clinical management protocol is several healthcare facilities worldwide (Naidu et al., 2022a).
Heparin in nebulized unfractionated form, provides a powerful anti-coagulant and mucolytic support to ameliorate respiratory symptoms, lower pulmonary dead space, and reduce ventilatory support in COVID-19 patients. Heparin is a promising prophylactic against VTE and could also relieve hypoxia-mediated symptoms in COVID-19 patients (Negri et al., 2020). Heparin-induced thrombocytopenia (HIT), a rare complication of heparin therapy, is estimated to occur in few patients. The repurposing of heparin and its derivatives as first-line therapy against SARS-CoV-2 is a promising strategy; however, this clinical approach needs further evaluation (Naidu et al., 2022a).
4.4. Iron (Fe)-chelators for nutritional management of COVID-19
Iron is an essential element for all living cells; however, free unbound iron from FeRD contributes to mitochondrial dysfunction, and dysbiosis of microbiota in lungs/gut (Edeas et al., 2020). Iron overload, free radical-induced tissue damage, thrombosis and erythrocyte dysfunction are implicated in hyperferritinemia, immune dysfunction and coagulopathy, a hallmark of severe COVID-19 (Muhoberac, 2020). Furthermore, free iron could target vascular tissues (i.e., hepatic, cardiac and endocrine cells), and cause severe damage to the corresponding organ function (Laforge et al., 2020). FeRD is a major cause of diffused endothelial inflammation with systemic involvement that could trigger an array of pathobiological manifestations during SARS-CoV-2 infection. Iron chelators inhibit IL-6 synthesis by down-regulation of NF-kB, and could suppress endothelial inflammation, a major risk factor for multi-organ failure in COVID-19 (Dalamaga et al., 2020). Furthermore, naturally occurring iron chelators, such as LF and transferrin (TF) could exert antiviral, anti-inflammatory and immunomodulatory effects that could be of high therapeutic value in the ongoing COVID-19 pandemic (Naidu et al, 2022b). Accordingly, iron chelators could play a potential role to ameliorate the systemic manifestations of COVID-19.
The FDA-approved oral chelators deferoxamine (DFO), deferiprone, and deferasirox can offer therapeutic solutions to treat iron overload and clinical conditions associated with free radical pathology (Kontoghiorghes and Kontoghiorghe, 2020). The natural siderophore DFO selectively removes iron from ferritin and hemosiderin to reduce the iron overload (Bellotti and Remelli, 2021. The potent antioxidant and free radical scavenging activities of DFO could be beneficial for highly vulnerable COVID-19 patients.
4.4.1. Phyto-chelators
Several plant-based compounds such as caffeic acid, curcumin, α-lipoic acid (ALA), and phytic acid are natural chelators that are known to protect cells from iron overload and restore mitochondrial membrane integrity, redox potential, and function. Caffeic acid, a plant-based iron-chelator, redox modulator, and a powerful natural antioxidant that could prevent lipid peroxidation in biological membranes (Hynes and O’Coinceanainn, 2004). Caffeic acid chelates could also interfere with viral attachment to heparan sulfate proteoglycans (HSPG) on cell surface (Langland et al., 2018). Curcumin from the Indian herb turmeric (Curcuma longa) is an iron-chelator that could inhibit iron-catalyzed pathways of oxidative stress and protect cellular DNA, lipids, and protein from free radical damage (Rainey et al., 2019). α-lipoic acid (ALA) could increase intracellular levels of glutathione (GSH), prevent the Nrf2 pathway activation during iron overload and restore mitochondrial membrane integrity, redox potential, and function (Camiolo et al., 2019). Phytic acid, abundant in edible legumes, cereals, and seeds, is an iron chelator with antioxidant activity could potentially inhibit iron-catalyzed hydroxyl (OH•) radical formation (Graf et al., 1987).
| 5. Post-acute sequelae of COVID-19 (PASC) | ▴Top |
Post-acute sequelae of COVID-19 (PASC), also referred to as the ‘long COVID’, has emerged as a novel clinical condition in COVID-19 survivors with lingering symptoms (or develop new ones) and fail to return to their baseline health. Accordingly, PASC is considered a post-COVID-19 sequelae with persistent symptoms and/or delayed or long-term complications beyond 4 weeks from the onset of acute or the initial phase of SARS-CoV-2 infection (Carfì et al., 2020; Nalbandian et al., 2021). Based on the duration of persistent clinical manifestations, PASC has been further categorized: i) post-acute COVID with symptoms and abnormalities that persist 4-12 weeks; and ii) chronic COVID with symptoms and abnormalities that persist >12 weeks and not attributable to alternative diagnoses (van Kampen et al., 2021; Shah et al., 2021). A recent cohort (based on 250,351 COVID-19 survivors from 2,100 studies), reported the high global prevalence of PASC: 54.0% at 1 month (short-term), 55.0% at 2 to 5 months (intermediate-term), and 54.0% at 6 or more months (long-term) (Groff et al., 2021). The burden of individual PASC sequelae vary by demography (age, race, and sex) but consistently higher among patients with existing metabolic syndromes and in survivors from severe acute infection (Xie et al., 2021).
The PASC is a multi-organ disorder ranging from mild symptoms to an incapacitating state and reduced quality of life that could last for weeks or longer following recovery from COVID-19 (Moghimi et al., 2021; Parker et al., 2021). The five most long-term clinical manifestations of PASC include fatigue (58%), headache (44%), attention disorder (27%), hair loss (25%), and dyspnea (24%), and generally have an impact on everyday functioning. (Lopez-Leon et al., 2021). Other common symptoms include shortness of breath, cough, joint pain, chest pain or tightness, loss of smell/taste, sore throat, diarrhea, depression, anxiety. Few less frequently observed symptoms include insomnia, palpitations, anorexia, tingling fingertips, and skin rashes (Ramakrishnan et al., 2021) (Figure 3).
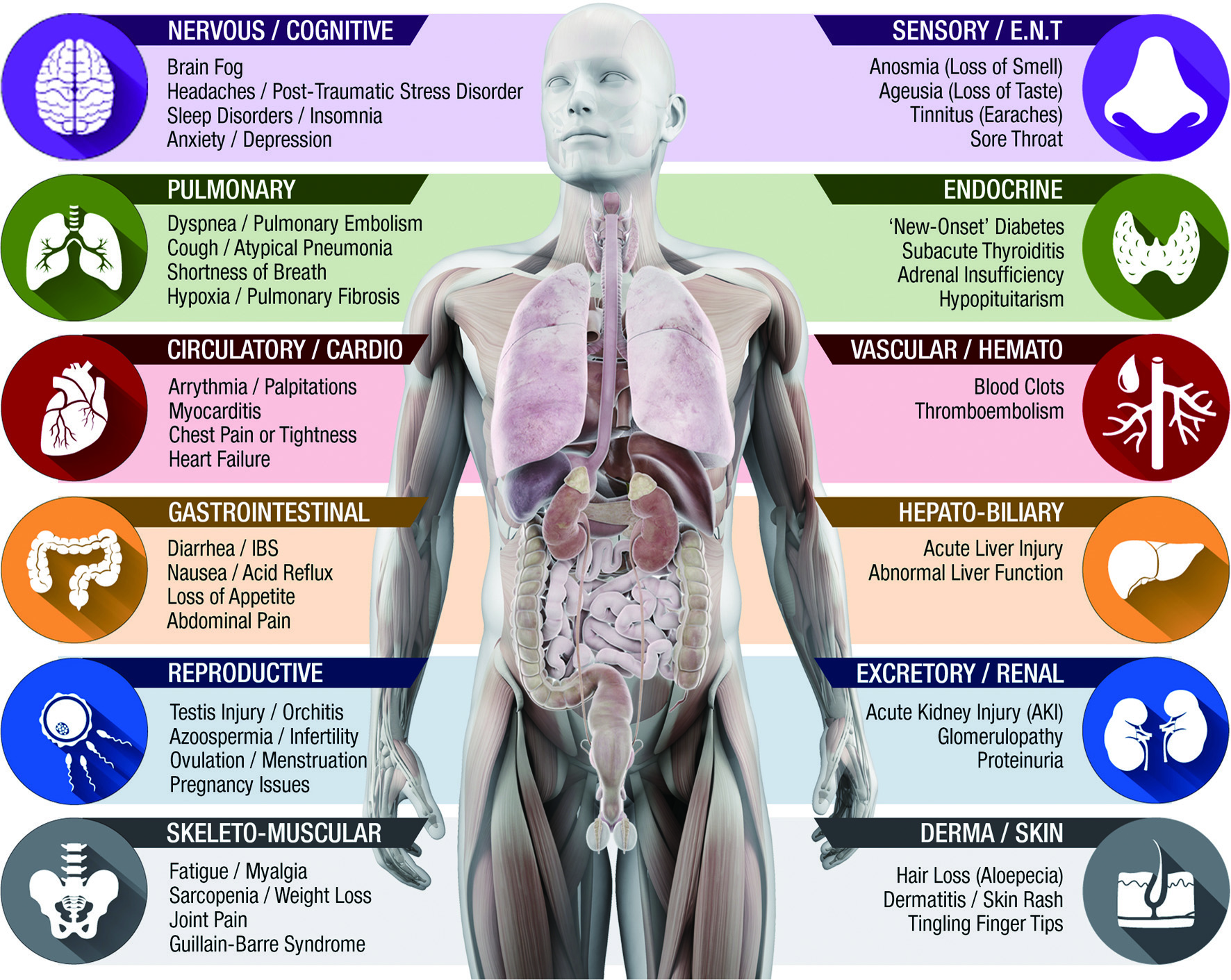 Click for large image | Figure 3. Post-acute sequelae of COVID-19 (PASC). Iron (Fe)-Redox Dysregulation (FeRD) Syndrome/host metabolic reprogram (HMR). |
5.1. Host metabolic dysfunction sequelae in PASC
The PASC pathology is a cumulative outcome of several viral-mediated alterations to host metabolism at cellular level such as: i) SARS-CoV-2 infection-induced tissue damage, ii) hyperinflammation-mediated multiorgan impairment, iii) immune exhaustion/dysregulation, iv) hormonal disturbances from maladaptation of ACE2-related pathways, v) coagulopathies due to endothelial damage/microvascular injury, iv) post-viral autoimmunity, vi) microbial dysbiosis, vii) critical care-associated sequelae or a combination of all above (Nalbandian et al., 2021; Moghimi et al., 2021; Ramakrishnan et al., 2021).
5.1.1. Dormant viral reservoirs
The prolonged symptoms of PASC may be attributed to earlier endothelial and tissue damage caused by SARS-CoV-2 during its acute clinical phases of infection. The multi-organ tropism of SARS-CoV-2 could also play a role in the lingering long-term sequelae of PASC, where the virus or viral epitopes may remain dormant and trigger chronic inflammatory responses. The dormant viral epitopes may also activate the adaptive immune cascade, which may lead to a persistent hyperinflammatory syndrome. The viral persistence and its probable genetic mutations could provoke antiviral ‘antibody waves’ leading to immune exhaustion which may further explain SARS-CoV-2 re-infections (Ramakrishnan et al., 2021).
5.1.2. Host immune exhaustion
The long-term sequelae of PASC suggests an accelerated rate of immune exhaustion in COVID-19 patients, a dysfunction of antigen-specific immune cells due to prolonged antigen stimulation (also from vaccination) (Diao et al., 2020; Zheng et al., 2020). Abnormal immune metabolism in may cause systemic perturbations such as FeRD, ROS/RNS production, oxidative and nitrosative stress (Naidu et al., 2022a).
Viral-induced autoimmunity is a possible contributing factor in the development of PASC. The SARS-CoV-2 virus may develop autoreactive T cells and antibodies to survive the post-acute infection or even develop post-viral clearance (Sette and Crotty, 2021).
Host mitochondrial dysfunction may predispose COVID-19 survivors to long-term health consequences and patients with metabolic syndromes such as diabetes and obesity are at higher risk of PASC (Scherer et al., 2022). Altered mitochondrial function and cellular energy deprivation leads to HMR with a metabolic switch from high energetic OXPHOS to low energetic glycolysis in COVID-19 patients (Singh et al., 2020b; Naidu et al., 2022a). Such patients demonstrate impaired cellular bioenergetics with altered metabolic pathways, including amino acids, lipids, sugars, and O2, using the limited energy reserves (Naviaux et al., 2016). A similar mechanism could be responsible for the chronic fatigue observed in PASC patients. In COVID-19 patients, the glycemic abnormalities could persist for at least 2 months after recovery, which suggests the impact of metabolic alterations in PASC pathobiology (Montefusco et al., 2021).
Altered host gut microbiome in COVID-19 patients, in tandem, with inflammatory and hematological markers, is an indicator of disease severity and dysfunctional immune response (Yeoh et al., 2021). The dysbiosis of gut microbiota persists for up to 30 days in COVID-19 survivors, which could play a critical role in the later onset of lingering PASC symptoms.
The renin-angiotensin system (RAS)/ACE2 axis plays a pivotal role in metabolic homeostasis and in the regulation of multi-organ functions in the body. ACE2, the enzyme critical for RAS activity, is also a host cell receptor for viral entry and the SARS-CoV-2 infection leads to a decline in ACE2 expression, subsequent increase in Ang II levels and potential hyper-activation of RAS (Ni et al., 2020). Elevated plasma Ang II levels have been associated with ALI/ARDS in severe cases of COVID-19 (Wu et al., 2020b). The imbalance between ACE2/angiotensin axis and RAS have also been implicated in multi-organ dysfunction syndrome (MODS) in COVID-19 (Rysz et al., 2021). The persistence of these clinical manifestations in post recovery suggests a significant involvement of ACE2 in PASC patients.
| 6. Nutritional strategies for PASC management | ▴Top |
PASC is characterized by malnutrition, loss of fat-free body mass, and low-grade inflammation. The recovery of these patients might be complicated by persistent symptoms such as functional impairment (i.e., fatigue and sarcopenia), dysphagia (particularly among intubated patients), loss of appetite, and taste/smell alterations (ageusia/anosmia) (Cereda et al., 2021). Therefore, any nutritional strategies for PASC management should address to rectify dietary deficiencies for adequate recovery of physical and functional wellbeing, as well as mental health of patients. The primary goal of such protocol is to prevent complications and support recovery to enable COVID-19 and PASC patients to achieve the best possible physical, functional, and mental health status.
6.1. Pulmonary impairments in PASC
Respiratory system is the most affected organ in COVID-19 and its clinical impact may extend into the post-COVID-19 phase even after patient recovery. In the respiratory sequelae, the SARS-CoV-2 invasion of alveolar cells leads to acute perivascular inflammatory response with severe tissue damage to pulmonary parenchyma (from vasculitis and endotheliitis), and to pulmonary vascular endothelium (due to huge infiltration of inflammatory cells) (Scholkmann and Nicholls, 2020). The ensuing breach of endothelial-epithelial barrier allows the infiltration of monocytes/neutrophils and extravasation of a protein-rich exudate into the alveolar space, consistent with other forms of ARDS (Huppert et al., 2019). The ARDS and atypical pneumonia could exert lasting damage to the lung alveoli through irreversible scarring or fibrosis in COVID-19 patients. This could lead to long-term breathing problems as well as the development of pulmonary fibrosis (Carsana et al., 2020). Pulmonary vascular micro-thrombosis and macro-thrombosis have been observed in 20–30% of patients with COVID-19 (Ackermann et al., 2020). The increased pulmonary microthrombi and macrothrombi formation in COVID-19 patients (Klok et al., 2020) may also contribute to the long-term respiratory PASC sequelae.
6.1.1. Pulmonary-PASC symptoms
Several COVID-19 survivors show long-term pulmonary post-discharge sequelae that include dyspnea (with or without chronic oxygen dependence), fatigue, impaired lung diffusion capacities, cough, and pulmonary fibrosis (Huang et al., 2021). Dyspnea is a common persistent symptom across varying degrees of initial COVID-19 severity and these survivors show reduced functional capacity and increased exertional desaturation of lungs (Cortés-Telles et al., 2021). Long-term risks of chronic pulmonary embolism and consequent pulmonary hypertension cannot be ruled out (Nalbandian et al., 2021).
6.1.2. Host metabolic targets for pulmonary re-optimization
Pulmonary impairment during COVID-19 is often accompanied by prolonged immobilization, which could compromise muscle function and lead to sarcopenia. Pulmonary rehabilitation should include nutrition, airway, posture, clearance technique, oxygen supplementation, breathing exercises, stretching, manual therapy, and physical activity to improve quality of life of PASC patients (Wang et al., 2020a).
6.1.3. Nutritional management of pulmonary-PASC
An analysis of 39 RCTs (n = 16,797) has identified various dietary supplements effective against viral respiratory tract infections (RTIs) (Shokri-Mashhadi et al., 2021). Vit-D could inhibit pulmonary inflammatory responses and enhance innate defense. Population-based studies showed a strong link between circulating Vit- D levels and lung function (Hughes and Norton, 2009). Vit-D improved RTIs across cohorts, particularly among Vit-D deficient patients. Specific Lactobacillus strains with selected prebiotics showed positive effects on the prevention and treatment of viral RTIs. A probiotic combination of Bifidobacterium sp. and Lactobacillus sp.,shown to reduce the symptoms of upper respiratory tract infection (Picó-Monllor et al., 2021). A supplementation with ginseng extract could also effectively prevent viral RTIs as adjuvant therapy. In a recent meta-analysis, Vit-C supplementation shown to reduce the risk of acute respiratory infection (ARI) and shorten the duration of symptoms. The effect of Vit-C on preventing ARI was stronger among men and in middle-income countries, compared to women and high-income countries, respectively (Abioye et al., 2021).
6.2. Neuro-cognitive impairments in PASC
Several neuro-COVID-19 impairments during PASC involve the central nervous system—CNS (i.e., central demyelination, seizures, encephalopathy/encephalitis, neurocognitive dysfunction, strokes); the peripheral nervous system—PNS (i.e., Guillain-Barréé syndrome/other neuropathies, neuralgias, myopathy, myositis), and the autonomous nervous system—ANS (i.e., dysautonomia, temperature and exercise intolerance), which increases long-term post-infectious disability (Thakur et al., 2021; Buoite Stella et al., 2022; Naidu et al., 2022c). The long-term neurological indications of COVID-19 could be attributed to direct viral infection, severe systemic and neuro-inflammation, microvascular thrombosis, and neurodegeneration (Heneka et al., 2020). The SARS-CoV-2 infection could damage brain parenchyma and vessels, possibly affect the blood-brain barrier (BBB)—cerebrospinal fluid barriers, which regulate neurons, supportive cells and brain vasculature (Reichard et al., 2020). Systemic/neuro-inflammation could lead to cognitive decline and the likelihood of neurodegeneration in COVID-19 survivors (Ramakrishnan et al., 2021). Furthermore, SARS-CoV-2 could affect the ANS and cause multiple neurological complications such as dysautonomia, orthostatic hypotension, and postural tachycardia syndrome (POTS) in PASC patients (Buoite Stella et al., 2022).
6.2.1. Neuro-PASC symptoms
Neurological complications in COVID-19 survivors frequent and represent a risk that compromises their functional capacity and the quality of life (Nordvig et al., 2021). More than half of COVID-19 survivors experience fatigue, apathy, executive deficits, impaired cognitive control, and reduction in global cognition; attributed to GABAergic impairment resulting from viral-induced neuro-inflammation (Ortelli et al., 2021). Neuromuscular manifestations such as dizziness, headache, myopathy, and olfactory and gustatory disturbances are frequently reported in PASC pathology (Shimohata, 2022). Migraine-like as well as late-onset headaches are prevalent PASC symptoms even after 6 weeks among 38% of post-discharged patients (Nalbandian et al., 2021). The underlying pathophysiology could be linked to the activation of peripheral trigeminal nerve endings by the SARS-CoV-2 directly or via vasculopathy and/or increased circulating pro-inflammatory cytokines and hypoxia (Bolay et al., 2020). Ageusia (loss of taste) and anosmia (loss of smell) resulting from olfactory dysfunction is a long-term PASC symptom reported persisted in more than 66% of European and US patients (Chiesa-Estomba et al., 2020; Garrigues et al., 2020).
Cough is another common PASC manifestations that lasts for weeks or months after SARS-CoV-2 infection. Such incessant post-COVID cough hypersensitivity state could be due to virus-mediated neurotropism, and neuroimmunomodulation of the vagal sensory nerves (Song et al., 2021b). Neuro-PASC patients demonstrate a specific immunological signature composed of humoral and cellular responses that are biased towards different SARS-CoV-2 structural proteins compared to healthy COVID convalescents, including a significant elevation in nucleocapsid (N)-specific antibody and T cell response (Visvabharathy et al., 2021).
6.2.2. Cognitive-PASC symptoms
Cognitive dysfunction, emotional distress, and functional decline are found to be prominent clinical symptoms in COVID-19 survivors at 4 months after acute infection (Vannorsdall et al., 2022). Psychological indications such as anxiety and depression; as well as mental health impairments such as delirium, ‘brain fog’, memory loss, hallucination, confusion, depression, and anxiety are long-term PASC manifestations (Rubin, 2020; Sudre et al., 2021). Post-COVID brain fog could result from dysautonomia, deconditioning or post-traumatic stress disorder (PTSD) (Kaseda and Levine, 2020). Long-term cognitive impairment has been reported in 20–40% of ICU discharged patients (Nalbandian et al., 2021). Also, COVID-19 seem to pose an increased risk of long-term cognitive decline in elderly population (Liu et al., 2021b).
6.2.3. Chemo-sensory dysfunction and PASC
Most COVID-19 patients (84.8%) show chemosensory dysfunction within the first 4 days of disease onset, and about 50% manifest this disorder in 2 to 3 weeks after infection. The chemosensory disturbance in terms of dysosmia and dysgeusia may persist in PASC patients (7.2%) even 60 days post-discharge (Vaira et al., 2020a). Dysosmia is a condition that affects perception of smell, which may lead to several chemosensory dysfunctional states including anosmia (total inability to detect odors), parosmia (altered and often displeasing odor perception), hyposmia (decreased ability to detect odors), and phantosmia (spontaneous odor detection without a trigger). Dysgeusia is a condition that affects perception of basic taste, which may lead to ageusia (total loss of the ability to taste) and parageusia (altered and often displeasing taste perception) (Hummel et al., 2011). On a long run, such debilitating olfactory and gustatory impairments could compromise the dietary intake of PASC patients with negative effects on their recovery. Therefore, nutritional strategies for COVID-19 and PASC management should consider appetite serving parameters to reduce malnutrition and support optimum patient recovery (Høier et al., 2021).
6.2.4. Host metabolic targets for neuro-cognitive re-optimization
PASC patients frequently report ‘brain fog’, a cognitive dysfunction involving memory problems, lack of mental clarity, and inability to focus. This dysfunction could be triggered by neuroinflammation from SARS-CoV-2 infection resulting in mast cell stimulation and release of microglial-activating mediators that inflame the hypothalamus (Marshall et al., 2019). Therefore, mast cell inhibition could be a potential therapeutic target to resolve brain fog-related issues during PASC.
6.2.5. Nutritional management of neuro-cognitive-PASC
Omega-3 polyunsaturated fatty acids (omega-3 or n-3 PUFAs) play a major role in immunity, inflammation, oxidative stress, and neurocognition at different symptomatic phases COVID-19. Omega-3 PUFAs, particularly EPA, is widely used to treat mood and neurocognitive disorders by reducing pro-inflammatory cytokines, altering the hypothalamus-pituitary-adrenal (HPA) axis, and modulating neurotransmission via lipid rafts. In addition, omega-3 PUFAs and their metabolites could ameliorate chronic inflammation, restore tissue homeostasis, and provide a promising strategy to treat ‘brain fog’ in PASC patients (Yang et al., 2022). Persistent inflammation, thrombosis, and a dysregulated immune response (auto-immune phenomena and/or persistent viral load) are major clinical manifestations of PASC. Oxidative stress and inflammation lead to development/progression of fatigue and neuro-psychiatric symptoms in several diseases by disrupting tissue integrity, blood flow and neurotransmitter metabolism. Intravenous Vit-C could help relieve these symptoms and reduce the risk of severe development of PASC (Vollbracht and Kraft, 2022).
Natural flavonoids, such as luteolin and quercetin are potential mast cell inhibitors that could ameliorate neuroinflammation and prevent cognitive dysfunction (Theoharides, 2020). Milk protein lactoferrin (LF) could effectively cross the blood-brain barrier and inhibits both microglia and mast cell-mediated inflammatory pathways in neuro-COVID-19 (Naidu et al., 2021c). Nutrients, including vitamins (B1, B6, B9, B12, C, D, and E), ω-3 fatty acids, and minerals (Fe3+, Zn2+, and Se2+), could help in down-regulation of neuroinflammation and oxidative stress and help recovery of PASC patients to regular cognitive state (Scarmeas et al., 2018; Motti et al., 2022). Oral intake of Vit-D has been suggested to prevent loss of neural sensation in COVID-19 patients by stimulating expression of neurotrophins such as the nerve growth factor (NGF) (Xu et al., 2020).
6.3. Cardiovascular impairments in PASC
The cardiovascular manifestations of COVID-19 19 may initially arise from subclinical pathology (i.e., myocarditis, pericarditis, palpitations, and right ventricular dysfunction) and ultimately evolve into myocarditis, stress cardiomyopathy, myocardial infarction, postural tachycardia syndrome (POTS), and arrhythmia (Chilazi et al., 2021). In most cases, myocardial injury is a direct outcome of COVID-19 severity, where ensuing myocardial fibrosis or scarring could manifest re-entrant arrhythmias (Liu et al., 2020d). Incidentally, the stress cardiomyopathy during the COVID-19 pandemic has been remarkably high (7.8%) compared to the pre-pandemic period (<2%), despite the unchanged CFR and re-hospitalization rates (Jabri et al., 2020). Based on the MRI data, the rate of myocardial inflammation seems to have increased as high as 60% after a 2-month diagnosis of COVID-19 (Puntmann et al., 2020).
6.3.1. Consequences of the viral spike (S) protein/ACE2 axis
The COVID-19 pathology perpetuates via the ACE2 receptor-mediated viral invasion of cardiac tissue (pericytes, cardiomyocytes, cardio-fibroblasts, epicardial adipose tissue, endothelia, and vascular cells), hyperinflammation, endothelial dysfunction with severe damage to myocardial/pericardial structural integrity and altered conduction system (Siripanthong et al., 2020). Such histopathological changes lead to cardiac sequelae (thromboembolism and blood pressure abnormalities) in PASC patients (Deshmukh et al., 2021; which are mediated by the dysregulation of RAAS and Kinin-Kallikrein System (KKS) (Cooper et al., 2021). Furthermore, COVID-19 survivors also demonstrate persistent cardio-metabolic demand with reduced cardiac reserve, and RAAS dysregulation.
ACE2, the type-I transmembrane metallo-carboxy-peptidase, is a critical regulator of the RAAS, and plays a vital role in the Fe-R-H status of cardiovascular and immune systems (Naidu et al., 2022a). The SARS-CoV-2 pathogen gains cellular entry is via the docking of the viral spike (S) protein to the membrane bound ACE2; and this infection process down-regulates ACE2 and/or sheds ACE2 from the cell surface (Cook and Ausiello, 2022). Such reduced ACE2 expression on cell surface could dysregulate Fe-R-H and initiate a plethora of cardiovascular impairments observed in COVID-19 and PASC (Chung et al., 2020).
6.3.2. Cardio-PASC symptoms
The cardiovascular system is affected not only during the acute phase of COVID-19, but also during the post-recovery phase. Cardiomyopathy could develop in post-COVID-19 patients due to persistent hyperinflammation, hypoxia, microvascular injury/thrombosis, coronary thrombotic/plaque rupture events, and direct viral cardiotoxicity from abnormal troponin levels (Sandoval et al., 2020). Fulminant myocarditis has been reported in several COVID-19 survivors even after weeks of undetectable viral pathogen (RT-PCR negative); however, some of these cases resulted in high fatality outcomes due to cardiac arrest (Inciardi et al., 2020)).
6.3.3. Nutritional management of cardiovascular-PASC
Dietary differences and ACE2 levels in populations influenced COVID-19-related CFR outcomes in several European countries. EU countries with high consumption of foods containing potent antioxidants or with anti-ACE activity (i.e., cabbage or fermented milk) showed low COVID-19-related mortality rate (Bousquet et al., 2020). Vit-C may be an effective treatment in decreasing the rates of mechanical ventilation and cardiac arrest in hospitalized patients with severe COVID-19 (Hess et al., 2022).
Cardiac injury is common manifestation associated with poor clinical outcomes in COVID-19 patients. In a retrospective cohort study, intravenous Vit-C (1.5 g/kg body weight) along with symptomatic supportive treatment to COVID-19 patients with ameliorated cardiac injury (ACI) group (n = 70), showed significant decrease in serum inflammatory markers (at day-21 during hospitalization). Therefore, Vit-C can ameliorate cardiac injury by alleviating hyperinflammation in severe and critically ill patients (Xia et al., 2021).
6.4. Renal impairments in PASC
Renal involvement in COVID-19 is frequent, which ranges from mild proteinuria to acute kidney injury (AKI). During the first COVID-19 wave, AKI was reported in nearly 1 in 3 COVID-19 patients and about 9% required kidney replacement therapy (KRT) (Lumlertgul et al., 2021). Viral-induced hyperinflammatory response and ischemic/hypoxic stress seems to be responsible for tubular, endothelial, and glomerular damage, a hallmark of septic AKI in COVID-19 (Diao et al., 2021; Long et al., 2022). Interestingly, several COVID-19 patients without any signs of AKI during the acute phase, show a gradual decline in renal function in 6–12-month follow-up period (Copur et al., 2022). The severity of COVID-19, older age, patients with comorbidities (diabetes, hypertension, and cardiovascular disease) are more prone to develop AKI (Yende and Parikh, 2021).
6.4.1. Renal-PASC symptoms
Renal sequelae leading to a progressive decline in kidney function has been widely reported in COVID-19 survivors. A decreased eGFR (estimated glomerular filtration rate) has been reported as a major indication in 35% of patients at 6-months in the post-acute COVID-19 (Huang et al., 2021). Collapsing glomerulopathy with involution of the glomerular tuft in addition to acute tubular injury (which is newly termed as the ‘COVID-19-associated nephropathy—COVAN’), may particularly impact patients of African ancestry, in some regions of the world (Velez et al., 2020). SARS-CoV-2-induced thrombotic microangiopathy with diffused cortical necrosis and microthrombi may also cause acute renal injury (Jhaveri et al., 2020).
6.4.2. Host metabolic targets for renal re-optimization
SARS-COV-2 infection could trigger the activation of multiple inflammatory pathways including angiotensin II, cytokine storm, C-reactive protein (CRP), TGF-β signaling, complement activation, and lung-kidney crosstalk could cause AKI. Hyper-inflammation plays a key role in the pathogenesis of AKI in patients; therefore, targeting these pathways with bioactive nutrients may represent a novel and specific dietary intervention for AKI resolution in PASC (Chen et al., 2021c).
6.4.3. Nutritional management of renal-PASC
In a meta-analysis (n = 1,459) a restricted protein diet supplemented with keto-analogs could effectively improve kidney endpoints including preserving kidney function and diminishing proteinuria, blood pressure levels, and CKD-mineral bone disorder parameters without causing malnutrition (Chewcharat et al., 2020). In another meta-analysis (8 RCTS/n = 371), supplementation with omega-3 fatty acid could decrease serum C-reactive protein levels in hemodialysis patients (Dezfouli et al., 2020). In another RCT (n = 60), patients with diabetes and chronic hemodialysis, supplemented with melatonin for 12 weeks showed beneficial effects on glycemic control and oxidative stress (Ostadmohammadi et al., 2020). In a clinical study, CKD patients (n = 28) given a diet supplemented with beta-glucan showed significant decline in trimethylamine N-oxide. Trimethylamine N-oxide levels, which is associated with severe kidney and cardiovascular outcomes (Hill et al., 2020). Quercetin, the natural anti-inflammatory agent, is shown prevent AKI and provide nephroprotective potential to COVID-19 patients (Wang et al., 2020b).
6.5. Gastrointestinal and hepato-biliary impairments in PASC
COVID-19 patients with gastrointestinal (GI) and hepato-biliary sequelae tend to experience severe clinical manifestations (i.e., ARDS and MODS) of SARS-CoV-2 infection (Dong et al., 2021). Gut impairment in COVID-19 patients has also been linked to the up-regulation of ACE2 and the ACE2 receptors in the GI tract (Hammoud et al., 2021). Prolonged fecal shedding of SARS-CoV-2 virus (RT-PCR positive) for >4 weeks after the onset of COVID-19 symptoms, as well as viral persistence for a mean of 11 d after negative RT-PCR of respiratory samples has been reported (Wu et al., 2020a). COVID-19 could alter the gut microbiome, including the enrichment of opportunistic infectious agents and depletion of beneficial commensals (Donati Zeppa et al., 2020). Intestinal microflora are known to influence the ‘gut-lung axis’ and could alter the course of respiratory infections (Bradley et al., 2019); accordingly, a butyrate-producing probiotic strain Faecalibacterium prausnitzii, associated with gut health was negatively correlated with the disease severity (Miquel et al., 2013; Zuo et al., 2020). Long-term consequences of COVID-19 on the GI tract, especially on post-infectious irritable bowel syndrome and dyspepsia is currently under clinical investigation (NCT04691895).
6.5.1. GI/hepato-PASC symptoms
Gastrointestinal (GI) sequelae such as diarrhea, nausea, acid reflux, loss of appetite, abdominal pain, and anorexia were observed in PASC patients at 90 days post discharge (Weng et al., 2021). In a systematic review (total studies: 43 / total patients: 18,246) diarrhea was reportedly common among 11.5% of the COVID-19 patients, followed by nausea and vomiting (6.3%) and abdominal pain (2.3%) (Silva et al., 2020). Also, abnormal liver function with increased ALT and AST levels was reported in 19% of PASC patients (Mao et al., 2020).
6.5.2. Host metabolic targets for GI re-optimization
SARS-CoV-2 could infect esophagus, stomach, duodenum, rectum, and the viral pathogen is detected in feces of COVID-19 patients. Prolonged clinical manifestation(s) of the virus in the GI, mainly the diarrhea, has been correlated with altered gut microbiota, immune dysregulation, and delayed SARS-CoV-2 clearance from the body. Elevated GI expression of ACE2 and TMPRSS2 proteins, the critical host cell factors for viral entry, could make gut epithelia a direct target for SARS-CoV-2 infection. Accordingly, the stool samples of COVID-19 patients exhibit proinflammatory cytokines (IL8), calprotectin (neutrophils activity), and IgA antibodies against the virus. Furthermore, the impairment of gut epithelial integrity could evoke hyper-immune response, hypoxia, and altered gut microbiota (dysbiosis) (Roy et al., 2021).
6.5.3. Nutritional management of GI and hepato-biliary-PASC
Medical care for patients hospitalized with COVID-19 is a challenging protocol. Most COVID-19 inpatients (58-95%) are commonly treated with broad-spectrum antibiotics to prevent ‘ventilator-associated pneumonia’ (VAP) and/or nosocomial infections. A meta-analysis (31 studies) showed that only 7% of hospitalized COVID-19 patients had bacterial co-infections; however, >90% received empirical antibiotics (Lansbury et al., 2020). Such wide use of antibiotics could pose a risk of ‘antibiotic-associated diarrhea’ (AAD) and Clostridium difficile infections (CDI) in some patients. About 20% of patients on antibiotics may contract AAD and the incidence varies depending on the type of antibiotic, age, co-morbidities and other risk factors (McFarland et al., 2016). Given the increase in antibiotic usage during this pandemic, there could be a possible resurgence of CDI-related complications in COVID-19 patients. Therefore, specific probiotic formulations could be promising adjuvants to combat both AAD and CDI in hospitalized COVID-19 patients. In a RCT, a multi-strain probiotic mixture was found to be effective in ameliorating COVID-19-associated diarrhea (Kullar et al., 2021). In a Chinese cohort study (n = 156), diarrhea was reported in 15.4% of COVID-19 patients and probiotic treatment seems to shorten the duration of diarrhea (Wang et al., 2021). The epithelial mucosa from respiratory and GI tracts are both affected from dysbiosis and inflammation; therefore, proper probiotic supplementation to re-establish healthy gut microbiota could be an important therapeutic strategy in the clinical management of COVID-19 and PASC.
6.6. Endocrine impairments in PASC
SARS-CoV-2 affects most endocrine glands including the pancreas, thyroid, and adrenal glands (Lundholm et al., 2020). The viral pathogen could affect endocrine system through various routes (i.e., direct viral injury, immunological and inflammatory damage) in emerging new-onset metabolic syndromes such as the development of type 1 diabetes mellitus, worsening of glycemic control (in pre-existing type 2 diabetes mellitus), primary Leydig cell damage, critical illness-related corticosteroid insufficiency, central hypocortisolism, pituitary apoplexy, immune-mediated hypophysitis, diabetes insipidus, sick-euthyroid syndrome, subacute thyroiditis, and bone demineralization with enhanced fracture risk (Pal and Banerjee, 2020; Makrydakis et al., 2022). These metabolic syndromes not only predispose the risk of severe COVID-19 but also increase the host susceptibility to SARS-CoV-2 infection and aggravate pre-existing endocrine disorders (Puig-Domingo et al., 2021). Recent studies have indicated that patients with history of diabetes have increased susceptibility to SARS-CoV-2 infection (Singh et al., 2020a; Abdi et al., 2020). Also, COVID-19 patients with underlying metabolic and vascular disorders have reportedly 50% case fatality rate (CFR) (Steenblock et al., 2021).
6.6.1. Endocrinal-PASC/new-onset metabolic syndromes
Disrupted hypothalamic-pituitary-thyroid (HPT) axis with abnormal thyroid function is a prominent endocrine manifestation in COVID-19 (Scappaticcio et al., 2021). The parathyroid dysfunction with hypocalcemia is reported in two-thirds of COVID-19 patients (Elkattawy et al., 2020). Both non-severe (1-2%) and severe (17%) COVID-19 patients show pancreatic injury (Liu et al., 2020c). Incidence of hyperglycemia in these patients is high (50%) (Ceriello, 2020), which is significantly associated with increased risk of mortality (Saand et al., 2021). Endocrinopathies in COVID-19 survivors include, hypopituitarism, central diabetes insipidus, SIADH, thyroid abnormalities, hyperglycemia, adrenal insufficiency, orchitis and alteration in sperm morphology (Mirza et al., 2022). Diabetic ketoacidosis (DKA) in COVID-19 survivors with no prior history of diabetes mellitus has been reported (Suwanwongse and Shabarek, 2021). Also, subacute thyroiditis with clinical thyrotoxicosis could ensue within weeks after the resolution of COVID-19 (Brancatella et al., 2020). SARS-CoV-2 infections could cause latent thyroid autoimmunity with new-onset Hashimoto’s thyroiditis186 or Graves’ disease (Mateu-Salat et al., 2020).
6.6.2. Host metabolic targets for endocrine re-optimization
ACE2 receptors are expressed in various endocrine tissues; thus, making the system a vulnerable target for SARS-CoV-2 infection (Lazartigues et al., 2020; Mirza et al., 2022). In endocrine system, ACE2 are localized in the paraventricular nucleus of hypothalamus (Chigr et al., 2020), in the acidophilic cells of parathyroid glands (He et al., 2006), in the pancreatic ductal, acinar and islet cells (Liu et al., 2020a), and co-expressed with TMPRSS2 in thyroid (Rotondi et al., 2021) and adrenocortical cells (Mao et al., 2021). Since ACE2 is an important component of the RAAS axis and a crucial entry point of SARS-CoV-2, the dynamics of ACE2 expression in endocrine tissue are also of contemporary relevance (Rath et al., 2021).
6.6.3. Nutritional management of endocrinal-PASC
Considering the pathobiological impact of endocrine disorders on COVID-19 pandemic, metabolic re-optimization with nutritional management should be considered an immediate priority. New-onset diabetes in PASC is not merely an extension of SARS-CoV-2 virulence but a combination of several host responsive intrinsic factors with chronic effects on host metabolism (Accili, 2021). Hyperglycemia-mediated aberrant glycosylation of ACE2 receptors could facilitates receptor-mediated viral invasion and increase the possibility of SARS-CoV-2 reinfection; therefore, glucose control with a strict dietary regimen an effective metabolic strategy for managing critically ill COVID-19 patients (Brufsky, 2020; Gianchandani et al., 2020).
The pathobiology of COVID-19 is closely associated with the host sirtuin activity—a family of NAD-histone deacetylase that regulate metabolic/redox homeostasis at cellular level with ameliorating effects on oxidative stress and inflammation (Huarachi Olivera and Lazarte Rivera, 2020; Wang and Wei, 2020). Sirtuin synthesis and activity decline with age, and its deficiency leads to an imbalance in redox homeostasis that could cause severe metabolic dysfunctions (Hall et al., 2013).
Dietary polyphenols such as pterostilbene and polydatin (precursors of resveratrol), are potential activators of sirtuins (Pacifici et al., 2019). Pterostilbene and polydatin can also have beneficial effects on metabolic diseases. In pre-clinical T2D, pterostilbene could ameliorate glycemic control, dyslipidemia, and liver injury (Zhang et al., 2020c). Oral administration of polydatin (50 mg/kg) in diabetic rats significantly enhanced glucose tolerance and insulin secretion (Yousef et al., 2021). Polydatin treatment could protect biological membranes from oxidative damage, preserve cell viability, and restore β-cell function.
6.7. Skeleto-muscular impairments in PASC
Skeleto-muscular impairments are common in both acute COVID-19 and PASC (Soares et al., 2022), which include sarcopenia, cachexia, myalgia, myositis, rhabdomyolysis, atrophy, peripheral neuropathy, and Guillain-Barré Syndrome. The risk of developing sarcopenia during COVID-19 illness or after recovery is relatively high (Seixas et al., 2022). In critically ill patients, the diaphragm damage has distinct myopathic features, which may contribute to long-term dyspnea and fatigue in COVID-19 survivors (Shi et al., 2021). Also, SARS-CoV-2 could affect the skeleto-muscular system indirectly via regulation of nerve impulses and blood supply. The viral interaction with ACE2 could dysregulate the RAAS activity and induce severe consequences such as loss of muscle mass, strength, physical dysfunction and delay the COVID-19 recovery process (Gonzalez et al., 2020). Elevated levels of IL6, the pro-inflammatory cytokine, could also trigger myalgia and joint pain (Roschel et al., 2020).
6.7.1. Skeleto-muscular-PASC symptoms
Myalgia (muscle pain)/fatigue is the third most common symptom (after fever, cough and sore throat) related to COVID-19 disease severity (Paliwal et al., 2020). Elderly COVID-19 patients with preexisting metabolic syndromes (i.e. diabetes, obesity, CVD) are highly prone to severe muscle injuries in later stages of COVID-19 (Pitscheider et al., 2021). Weight loss is common usually affecting non-fat mass (especially in obese patients), which may inflict dystrophic damages to skeletal muscle in COVID-19 survivors (Haraj et al., 2021). One in five hospital admitted COVID-19 patients show serious weight loss and 73% experience high risk of sarcopenia (Wierdsma et al., 2021). Rhabdomyolysis, an acute muscle injury with intense muscle soreness, fatigue, weakness, and lower limb pain/twitching, often observed in elderly patients, is a characteristic feature of severe COVID-19 (Jin and Tong, 2020). COVID-19-related sarcopenia and rhabdomyolysis could lead to long-term disabilities.
6.7.2. Host metabolic targets for skeleton-muscular re-optimization
Skeletal muscle is the largest body tissue involved in glucose metabolism (Riuzzi et al., 2018); therefore, a primary target for SARS-CoV-2 infection (Ali and Kunugi, 2021a). The viral-induced hyper-inflammatory response may also exacerbate mitochondrial dysfunction with subsequent myofibrillar breakdown and muscle degradation (Piotrowicz et al., 2021). Elevated synthesis of creatine kinase (CK), lactate dehydrogenase (LDH), and myoglobin (a heme-containing globular protein) in hypercatabolic conditions trigger oxidative stress and leads to severe myocyte damage in COVID-19 (Welch et al., 2020). The decline in muscle mass (sarcopenia) is a whole-body process that also affects respiratory, masticatory, and swallowing functions, which may also have a negative impact on nutrient intake and increase the risk for malnutrition. In a cohort, more than 90% of COVID-19 patients showed some degree of dysphagia, with the need of a modified diet consistency or nasogastric feeding (Brugliera et al., 2020).
6.7.3. Nutritional management of skeleto-muscular-PASC
Several COVID-19 survivors may have sequelae of muscle wasting and require progressive dietary plan for gradual recovery to pre-onset mobility function. Malnutrition has been attributed for muscular and immune dysfunction in COVID-19 (Ali and Kunugi, 2021b). Age-related sarcopenia, similar to muscle loss in COVID-19, could be effectively reversed by high protein supplementation (Liao et al., 2019). Accordingly, effective nutritional strategies (i.e., protein-rich diet, and specific bioactive food supplements) could minimize the risk of sarcopenia in vulnerable COVID-19 patients (Chapple et al., 2020; Cawood et al., 2020). Specific marine protein hydrolysates are shown to interact with SARS-CoV-2 enzymes (Mpro and monoamine oxidase A) and may reduce the viral load and associated disease severity (Yao et al., 2020). Oral administration of L-glutamine could shorten the hospital stay and lower the CFR in COVID-19 patients (Cengiz et al., 2020). Also, exposure to sunlight could stimulate vitamin D synthesis and promote muscle protein synthesis as well as strengthen the immune system (Xu et al., 2020). Specific nutritional regimen (i.e., protein diet and amino acid supplements) with adequate physical exercise may help restore skeletal muscle metabolism and avert the after-effects of physical disability in COVID-19 survivors.
Hyper-inflammatory state during COVID-19 could negatively affect the muscle protein synthesis and this progressive disorder leads to severe catabolic wasting of skeletal muscle mass and function loss, a condition known as sarcopenia (Crispo et al., 2021). Acute sarcopenia is prevalent among elderly COVID-19 patients, with direct implications in PASC onset and recovery (Piotrowicz et al., 2021). Vit-D supplementation could help in muscle recovery after damage (i.e., sarcopenia) from SARS-CoV-2 infection. Vit-D supplementation (cholecalciferol: 2,000 IU/d) could decrease serum creatinine kinase levels to optimum values, improve the health status and quality of life of COVID-19 patients during the recovery process (Caballero-García et al., 2021). Nutritional support to restore muscle mass is critical in the management of PASC. The minimum protein requirement is about 0.83 g/kg body weight per day for elderly subjects. In addition, supplementation with leucine, β-hydroxy-β-methyl-butyrate or creatine could be effective (Gielen et al., 2021). Such supplemental support (or in the form of a balanced diet) may help support muscle mass restoration in PASC patients with post-COVID-19 syndrome.
6.8. Reproductive impairments in PASC
The hypothalamic-pituitary-adrenal (HPA), -thyroid (HPT), and -gonad (HPG) axes are readily impaired during SARS-CoV-2 infection, which may lead to sexual dysfunction in males (Pal, 2020). In general, the testis is shielded from external factors by the blood-testicular barrier; however, SARS-CoV-2 could easily breach this physiological hurdle and cause inflammation (Massarotti et al., 2021). Host receptor (ACE2) and serine protease (TMPRSS), both critical for the viral cellular entry, are widely co-expressed in the testes, which makes the male reproductive system a potential target for SARS-CoV-2 infection (Zupin et al., 2020). ACE2 is also abundant in gametocytes, Sertoli cells and spermatogonia stem cells, that makes testis a high-risk organ in COVID-19 (Vishvkarma and Rajender, 2020). Thus, male gonads may be potentially vulnerable to SARS-CoV-2 infection; therefore, caution should be exercised by expectant women and couples planning natural pregnancy or assisted reproduction.
6.8.1. Male reproductive complications
Male patients with COVID-19 demonstrate elevated levels of prolactin and luteinizing hormone (LH) in contrast to low testosterone and follicle stimulating hormone (FSH), which indicates primary testicular damage during active illness (Selek et al., 2021). However, several COVID-19 patients exhibit damaged seminiferous tubules, reduced Leydig cell number, swollen Sertoli cells and mild lymphocytic inflammation in the testes (Yang et al., 2020a). Severe damage to seminiferous tubules with interstitial edema, congestion, RBC exudation in testes, and epididymis, indicate a potential testis injury in COVID-19. Furthermore, semen specimens from most patients (39%) indicate oligo-zoospermia (increased seminal levels of IL-6, TNF-α, and MCP-1) with impaired spermatogenesis (Li et al., 2020b). These patients also show scrotal discomfort, which is suggestive of viral orchitis. Also, a number of patients (31%) showed significant alterations in semen parameters including sperm DNA fragmentation and reduced sex-related hormone ratios, i.e., testosterone to luteinizing hormone (T:LH) and FSH:LH (Ma et al., 2021a). Furthermore, SARS-CoV-2 has also been detected in the semen of males with acute infection as well as in recovering patients (Polack et al., 2020). These results collectively suggest that COVID-19 may lead to potential male fertility issues. Therefore, COVID-19 survivors especially individuals in reproductive age, andrological consultation and evaluation of gonadal function including semen analysis has been highly recommended (Corona et al., 2020).
6.8.2. Female reproductive complications
The SARS-CoV-2 could also invade the female reproductive system through the ACE2/TMPRSS2-mediated host viral entry mechanism. The ACE2 receptor is widely expressed in the ovary, uterus, vagina, and placenta, which in tandem with Ang II and Ang-(1-7) regulates the follicle development/ovulation, modulate luteal angiogenesis/degeneration, influence the regular changes in endometrial tissue and embryo development in the female reproductive tract (Jing et al., 2020); therefore, COVID-19 could possibly affect ovulation and pregnancy. There is an abundance of ACE-2 receptors in the ovaries and oocytes; however, the existing data on possible ovarian dysfunction in COVID-19 survivors or any long-term sequelae on female fertility needs further investigaton (Segars et al., 2020; Stanley et al., 2020).
6.8.3. Host metabolic targets for reproductive re-optimization
Both male and female COVID patients could develop sexual and reproductive health issues on the long run, partly due to the ACE2/TMPRSS2 axis that enables SARS−COV-2 entry into either reproductive systems (i.e., male gonads or female ovaries) (Omolaoye et al., 2021). Interestingly, ACE 2 is highly expressed in the testes of infertile men than normal, which suggests that COVID-19 may cause male reproductive disorders via ACE2 activated pathway and men with reproductive issues may be more susceptible to SARS-CoV-2 infection (Zupin et al., 2020). The degree of ACE2 expression is age-related; accordingly, the middled-aged have demonstrated higher incidence of SARS-CoV-2 infection compared to young men (Shen et al., 2020). Several postulated mechanisms include Fe-R-H dysregulation, inflammation-induced oxidative stress, HPA dysfunction and direct gonadal damage (Dutta and Sengupta, 2021; Seymen, 2021; Mannur et al., 2021).
6.8.4. Nutritional management of reproductive-PASC
Adequate maternal Se2+ levels are vital for immune response and healthy pregnancy. Serum Se2+ levels of pregnant women in the 2nd and 3rd trimesters with COVID-19 (n = 71) were significantly lower than in the healthy group (n = 70). Serum Se2+ levels gradually drop during pregnancy; however, this natural decline is enhanced with COVID-19 infection. The drop in maternal Se2+ levels correlated with elevated IL-6 and D-dimers, which indicated the effect of selenium on COVID-19 progression (Erol et al., 2021). SARS-CoV-2 infections are more prevalent in men than women, partly due to higher expression of ACE2 in the male reproductive tissues (Zupin et al., 2020). The seminal dissemination of SARS-CoV-2 and its mediated testicular disruptions have been reported. Vit-C is a major testicular antioxidant that neutralizes excess ROS, prevents sperm agglutination, blocks lipid peroxidation, recycles Vit-E, and protects testicular tissue against DNA damage. Thus, Vit-C could be an effective intervention to alleviate testicular oxidative stress and associated male reproductive dysfunctions in COVID-19 and PASC (Sengupta et al., 2022).
Balanced diet could provide essential nutrients, confer adequate nutritional status, and improve metabolic activity of a COVID-19 or PASC patient to recover/regain optimal health. The nutritional status of a patient is critical for effective outcomes of antiviral drugs, vaccines, and other therapeutic regimens used in the clinical management of COVID-19 and PASC. An adequate dietary plan should (i) reduce post-viral fatigue, (ii) promote earliest recovery, and (ii) provide future resistance in often malnourished patients (i.e., elderly, obese, diabetics, etc) (Butters and Whitehouse, 2021).
Nutritional status of the patient population has been severely compromised (directly or indirectly) by the COVID-19 pandemic. Nutritional interventions through dietary support, food supplementation, and natural remedies could help manage PASC, the aftermath of COVID-19 survival. The nutrition-COVID-19 interaction combined with the altered dietary intakes of COVID-19 survivors may lead to malnutrition and the inherent micronutrient deficiencies resulting in metabolic disorders (i.e., new onset diabetes), symptomatic progression to PASC with potential morbidity and mortality scenarios (Antwi et al., 2021).
Nutrition could play a vital role in PASC management. Several food bioactives are known to deliver multifunctional benefits to different host cellular targets, help reduce symptomatic burden and revitalize affected organ/systems, through independent as well as synergistic mechanisms. Nutritional inadequacy during hospitalization exacerbates the likelihood of long-term PASC syndrome. Malnutrition could delay healing and increase the duration of hospitalization. Monitoring the nutritional status is critical in the management of hospitalized COVID-19 patients and in the rehabilitation of PASC individuals. Therefore, an early dietary support with oral nutritional supplements (ONS), enteral and parenteral nutrition, may ameliorate such detrimental effects and play a vital role in clinical management as well as aiding the recovery of hospitalized patients (Volkert et al., 2019). The European Society for Parenteral and Enteral Nutrition (ESPEN) recommends that COVID-19 patients should be given ONSs that provide at least 400 kcal/day and ≥30 g protein/day (Barazzoni et al., 2020).
| 7. Nutritional strategies for COVID-19 management: a global health perspective | ▴Top |
Malnutrition (undernutrition) is highly prevalent among COVID-19 patients, which is about 37% in general patients, 52.7% among elderly patients and 66.7% among patients admitted in ICU (Latif et al., 2022). Duration of hospital stay for malnourished patients infected with SARS-CoV-2 is significantly higher (almost double) than the non-malnourished cases (Yu et al., 2021). The SARS-CoV-2 infection could severely compromise the dietary intakes of patients in several ways (Holdoway, 2020). The pulmonary-related COVID-19 issues such as coughing and breathlessness, dry mouth due to oral breathing, use of inhalers and oxygen therapy, may cumulatively affect the eating and drinking regimen of COVID-19 patients (British Lung Foundation (BLF), 2020). Also, the sensory loss of smell (anosmia) or taste (ageusia)—the distinctive symptoms of COVID-19, could affect patient’s appetite and desire to eat (Xydakis et al., 2020). A raise in body temperature may increase the nutritional demand; however, the ensuing hyper-inflammatory response could suppress appetite and lead to muscle catabolism (Gandy, 2015). Furthermore, the aging process profoundly alters several physiological functions in the body, that eventually decreases the body’s ability to cope with stressors. Many such changes are subtle but could negatively impact nutrient intake and lead to malnutrition.
The pathobiology of COVID-19 could negatively impact the metabolic parameters of a patient at different levels such as: i) by increasing specific nutritional demand as a consequence of pyrexia, sepsis, dyspnea, and/or ii) by reducing dietary intake due to excessive coughing, dysphagia, dysgeusia, chronic fatigue, poor appetite and food access issues (Barazzoni et al., 2020). Therefore, nutritional strategies or recommendations should consider both the viral-specific virulence targets (i.e., viral entry blockers and viral replication inhibitors), as well as specific host defense factors (i.e., anti-inflammatories, immune modulators, antioxidants, and metabolic optimizers) for obtaining effective outcomes. The following section elucidates such target-specific nutritional strategies for COVID-19 management (Figure 4).
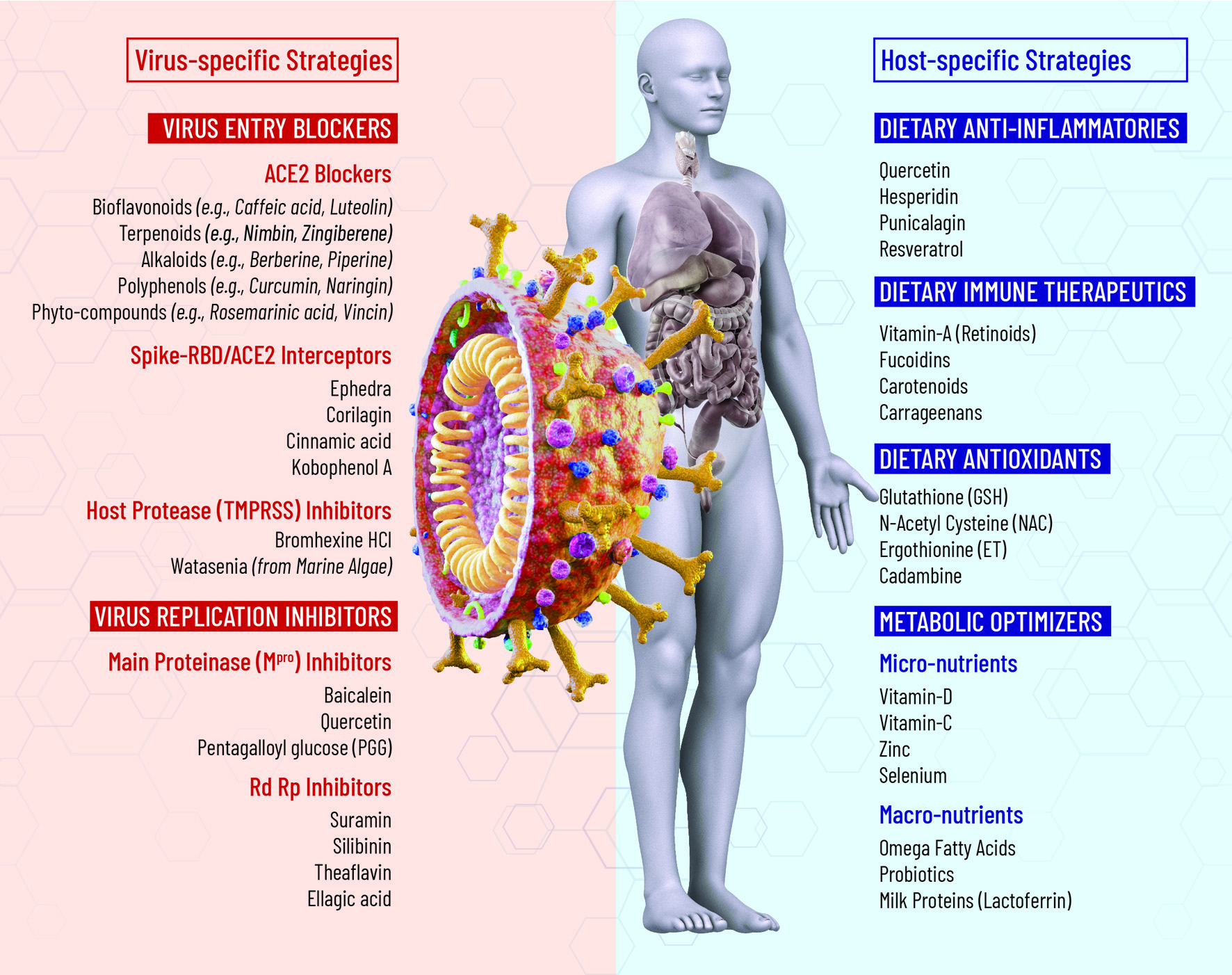 Click for large image | Figure 4. Target-specific nutritional strategies for COVID-19 management. |
7.1. Phytochemical blockers of viral-host interaction
Despite global immunization campaigns, the prophylactic efficacy of vaccines seems to be compromised due to emerging ‘variants of concern’ (VOC). Therefore, development of host-directed therapeutics and prophylactics could contain such resistance and offer some urgently needed protection against VOCs. Antiviral drugs with proven efficacy are not yet available to prevent transmission or facilitate treatment of COVID-19. Natural compounds, especially plant-derived bioactives, have emerged adjuvant interventional options to overcome the limitations of existing antiviral drugs against COVID-19 (Patil et al., 2021). In the onoing COVID-19 pandemic, several clinical practices have integrated complementary or traditional medicine, as adjuvant therapeutic protocol with the Western medicine (Wan et al., 2020; Mani et al., 2020).
Computational drug repurposing is a promising alternative that enables prioritization of existing compounds through rapid high through-put screening analyses (Harrison, 2020). Several ongoing in silico studies have demonstrated the antiviral potential of natural compounds and these phytochemicals also have multifunctional effects such as anti-inflammatory, antiviral, antioxidant, cardioprotective, and exhibit potent therapeutic benefits in the treatment of COVID-19 associated clinical manifestations. The ‘repurposing’ of approved antiviral drugs with adjuvant combination(s) of well characterized phytochemicals could be one of the rapid and safe strategies to combat the COVID-19 pandemic (Naidu et al., 2021a; 2021b).
7.1.1. Host receptor (ACE2) blockers
Angiotensin-converting enzyme 2 (ACE2), also known as peptidyl-dipeptidase A, is a type I integral membrane protein. ACE2 has been implicated in the regulation of heart function and also as a functional receptor for the SARS-CoV-2 pathogen. The interaction between SARS-CoV-2 Spike (S)-protein and the host ACE2 receptor is essential for viral attachment and the subsequent fusion process. The interaction between the SARS-CoV-2 S-protein and ACE2, triggers activation of several host membrane molecules such as the enzyme A disintegrin and metalloproteinase 17 (ADAM17) that regulates ACE2 expression, and transmembrane protease serine 2 (TMPRSS2) that facilitates virus-cell membrane fusion (Zlacká et al., 2021). Blocking the activity of these host membrane factors is a promising strategy in the development of prophylactic and therapeutic for COVID-19 management. Accordingly, ACE2 has emerged as a potential antiviral intervention target against SARS-CoV-2.
Most ACE2 blockers in practice are synthetic chemicals with limitations in molecular stability and adverse side effects. However, natural, and selective ACE2 blockers are highly stable with excellent safety/tolerance profiles and could substitute the synthetic drug inhibitors (Pokhrel et al., 2021). Historically, plants served as bio-factories for the isolation of various bioactive compounds with multifunctional health benefits and provided multitude of possibilities for therapeutic interventions. Molecular docking and ‘molecular dynamics simulation’ (MDS) studies have aided in rapid screening of molecular interactions between effective phytochemical ligands (ACE2 blockers) and the receptor binding domain (RBD) of viral S-protein. This could provide strategies to identify effective S protein-ACE2 blockers from natural sources and explore the possible development of broad-spectrum anti-COVID interventions. (Ma et al., 2021b) To combat the current COVID-19 pandemic, several plant-based natural compounds such as flavonoids, terpenoids, phenols, alkaloids, have been identified for their potential to inhibit the SARS-CoV-2 pathogen and/or reinforce host immune defenses.
7.1.1.1. Phytochemical interactions with ACE2 protein
Several flavonoids are known to inhibit ACE2 activity and play a vital role in the regulation of arterial blood pressure (Muchtaridi et al., 2020). Bioflavonoids from ethanolic extracts of bee propolis—caffeic acid, caffeic acid phenethyl ester, chrysin, galangin, myricetin, rutin, hesperetin, pinocembrin, luteolin and quercetin demonstrate high-affinity binding to ACE2 receptors (Guler et al., 2021). Five compounds from sulawesi propolis—Tetragonula sapiens: glyasperin A, broussoflavonol F, sulabiroins A, (2S)-5,7-dihydroxy-4′-methoxy-8-prenylflavanone and isorhamnetin strongly inhibited the interaction of ACE-2 with SARS-CoV-2 (Khayrani et al, 2021). Several terpenoids (i.e., nimbin, withaferin A, andrographolide, zingiberene) and alkaloids (i.e., berberine, piperine, and thebaine) show high-affinity binding to ACE2 target (in silico). Also, phyto-phenolic compounds, including quercetin, curcumin, naringenin, luteolin, hesperidin, mangiferin, and gallic acid could avidly bind to ACE-2 (in silico), and the bioactive esculetin could decrease ACE-2 expression (in vivo) (Junior et al., 2021). Natural compounds such as hinokinin from grapes (Vitis vinifera), gmelanone from ‘gamhar’ (Gmelina arborea), isocolumbin, and tinocordioside from ‘giloy’ (Tinospora cordifolia) as potent ACE2 inhibitors, which may block SARS-CoV-2 infection (Sharma et al., 2021). Phytochemicals from crofton weed (Ageratina adenophora), 5-β-glucosyl-7-demethoxy-encecalin (5GDE) and 2-oxocadinan-3,6(11)-dien-12,7-olide (BODO) were found to be potential blockers of ACE2 (Neupane et al., 2021). Phytoconstituents of ‘long pepper’ (Piper longum) and ‘holy basil’ (Ocimum sanctum)—vicenin 2, rosmarinic acid, and orientin could modulate ACE2 and TMPRSS2 expression in host cells and may therapeutic potential against COVID-19 (Jindal and Rani, 2022).
7.1.1.2. Spike (S)-RBD/ACE2 binding interceptors from medicinal herbs
Ethanol extracts from the Traditional Chinese Medicine (TCM) herb—‘ephedra’ (Ephedra sinica) could effectively intercept the interaction between ACE2 and SARS-CoV-2 spike protein receptor-binding domain (RBD) (IC50 = 95.01 μg/mL) (Mei et al., 2021). Three active compounds, 4,6-dihydroxyquinoline-2-carboxylic acid (IC50 = 0.58 μM), 4-hydroxyquinoline-2-carboxylic acid (0.07 μM), and 4-hydroxy-6-methoxyquinoline-2-carboxylic acid (0.15 μM) were identified to intercept ACE2-RBD interaction. These findings suggested that quinoline-2-carboxylic acids from ephedra could be potential phyto-interceptors for COVID-19. Extracts from ‘danshen’ (Salvia miltiorrhiza Bunge), widely used in TCM, could effectively intercept SARS-CoV-2 binding to ACE2 receptor, as well as mitigate the inflammatory responses by interfering with the NFκB signaling (Petitjean et al., 2022). Geraniin, the main ingredient of the medicinal plant Elaeocarpus sylvestris var. ellipticus and Nephelium lappaceum, could effectively block the binding of viral S-protein with hACE2 receptor, suggesting that geraniin might inhibit the entry of SARS-CoV-2 into human epithelial cells (Kim et al., 2021). The extract from Nigella sativa seed (black seed oil), β-bisabolene showed a high-affinity binding to the ACE2 target (- 8.0 kcal/mol) almost similar to the antiviral drug Remdesivir (- 8.1 kcal/mol) (Duru et al., 2021). Bioactive extracts the neem (Azadirachta indica), azadirachtin H, quentin and margocin were identified as potential viral cell entry inhibitors with ability to effectively block SARS-CoV-2 spike RBD-ACE2 interaction (Shadrack et al., 2021). Isothymol derived from the essential oil of Ammoides verticillate is a functional inhibitor of ACE2 activity with best docking scores, compared to Captropil (ACE2 inhibitor drug) and Chloroquine (Abdelli et al., 2021).
In a recent study, N-0385, a small-molecular compound, has demonstrated low nanomolar potency and could inhibit SARS-CoV-2 infection in human lung epithelia and in donor-derived colonoids (Shapira et al., 2022). This compound also inhibited the cellular (Calu-3 cell model) entry of SARS-CoV-2 VOCs B.1.1.7 (Alpha), B.1.351 (Beta), P.1 (Gamma) and B.1.617.2 (Delta). Notably, in the K18-human ACE2 transgenic mouse model of severe COVID-19, the N-0385 has elicited a high level of prophylactic and therapeutic benefit after multiple administrations or even after a single administration. The N-0385 could provide an effective early intervention strategy against COVID-19 and emerging SARS-CoV-2 variants of concern.
7.1.1.3. Natural phyto-nutrient interceptors of spike (S)-RBD/hACE2 interaction
Phytochemical extracts such as cinnamic acid (-5.20 kcal/mol), thymoquinone (-4.71 kcal/mol), and andrographolide (-4.00 kcal/mol) potentially intercept interactions between ACE2 and the SARS-CoV-2 receptor protein. These compounds could efficiently activate the biological pathway without changing the conformation in the binding site of the ACE2 receptor (Srivastava et al., 2021). Naturally occurring triterpenoids such as glycyrrhetinic and oleanolic acids, as well as primary and secondary bile acids and their amidated derivatives such as glyco-ursodeoxycholic acid and semi-synthetic derivatives such as obeticholic acid could potentially interfere in RBD/ACE2 binding and serve as phytochemical SARS-CoV-2 entry inhibitors (Carino et al., 2020). The natural polyphenol—Kobophenol A, could intercept the interaction of viral spike (S1)-RBD domain with ACE2 receptor (IC50 = 1.8 μM), as well as inhibit SARS-CoV-2 infection of host cells (EC50 = 71.6 μM) (Gangadevi et al., 2021). Natural stilbenoids analog, hopeaphenol (a resveratrol tetramer, IC50 = 0.11 μM), vatalbinoside A and vaticanol B, are potent and selective interceptors of RBD/ACE2 binding and viral entry across multiple VOCs (Tietjen et al., 2021). Natural compounds (dosage:100 μM)—epigallocatechin gallate (EGCG), ginsenoside Rg3, isobavachalcone, isochlorogenic A and bakuchiol, effectively intercept pseudo-virus entry, inhibit SARS-CoV-2-induced cytopathic effect and plaque formation in Vero E6 cells. Interestingly, these compounds bind to ACE2 receptor that suggests a dual interaction with both RBD and ACE2 (Zhang et al., 2021a).
A novel mannose-binding plant lectin from Narcissus tazetta bulb, NTL-125, could effectively intercept SARS-CoV-2 replication. In silico docking studies revealed that NTL-125 has strong affinity to spike (S)-RBD, intercept the viral attachment to hACE2 receptor, the gateway to cellular entry of the pathogen (Sarkar et al., 2022). The unique α-helical tail of NTL-125 plays most important role in binding to S-RBD. NTL-125 also interacts with certain glycan moieties of S-protein in addition to amino acid residues adding to the binding strength. Thus, NTL-125 is a highly potent antiviral compound of natural origin for management of COVID-19.
Corilagin derived from ‘leaf flower’ (Phyllanthus urinaria) demonstrate high-affinity interaction with SARS-CoV-2-RBD or human ACE2 (hACE2) protein (Yang et al., 2021b). Corilagin intercepts SARS-CoV-2-RBD binding in a dose-dependent manner and abolish the infectious property of RBD-pseudo-typed lentivirus, which mimics the entry of SARS-CoV-2 virus in human host cells. Corilagin (300 mg/kg/day) is safe and could be a potential antiviral agent against the COVID-19 that blocks the fusion of SARS-CoV-2 spike-RBD to hACE2 receptors. Natural polyphenols 1,3,6-tri-O-galloy-β-d-glucose (TGG) and, to a lesser extent, corilagin, could block the SARS-CoV-2 viral entry into target cells via the binding of RBD to ACE2 (Binette et al., 2021).
7.1.2. Host protease (TMPRSS) inhibitors
For host cellular entry, the surface spike (S)-protein of SARS-CoV-2 must be cleaved at two different sites by host cell proteases, the ‘proprotein convertase furin’ at the S1/S2 site and the transmembrane serine protease 2 (TMPRSS2) at the S2’ site (Bestle et al., 2020). Type-II transmembrane serine proteases (TTSPs) such as TMPRSS2; that cleave the viral spike (S)-protein to expose the fusion peptide for cell entry (Hoffmann et al., 2020). The TTSP-mediated proteolytic maturation of the S-protein is critical for SARS-CoV-2 infection and any host-directed targets to impede this viral entry mechanism could provide a potential antiviral intervention. Although the cellular entry of SARS-CoV-2 depends on ACE2, the virus also needs TMPRSS2 for its spike protein priming. It has been shown that the entrance of SARS-CoV-2 through ACE2 can be abrogated by cellular TMPRSS2 inhibitors.
Several TMPRSS2 inhibitors—camostat mesylate, nafamostat mesylate, antiandrogens, inhaled corticosteroids and ADAM-17 enhancers (5-fluorouracil) are already in clinical practice (Ragia and Manolopoulos, 2020). Bromhexine hydrochloride, the FDA-approved expectorant/mucolytic agent/cough suppressant, is a potent inhibitor of TMPRSS2 (Shen et al., 2017). Therefore, blocking of pulmonary TMPRSS2 with bromhexine hydrochloride could represent a prophylactic strategy against the airborne transmission of SARS-CoV2 (Habtemariam et al., 2020). Marine natural bioactive, Watasenia β-D-Preluciferyl glucopyrasoiuronic acid, is a potent inhibitor of TMPRSS2 with exhibited favorable drug-like pharmacokinetic properties to combat SARS-CoV-2 (Mahmudpour et al., 2021).
7.2. Phytochemical inhibitors of viral replication
Natural compounds and bioactives from medicinal herbs that are widely used for centuries in global healthcare, could pave a new direction in the development of novel anti-COVID-19 prophylactics and therapeutics (Mahrosh and Mustafa, 2021a). Interestingly, many potential drugs in current practice are based on the structure of natural compounds with desired biological activities. Almost half the drugs approved between 1981 and 2014 by the US-FDA, were derived from or mimicked a natural compound (Newman and Cragg, 2016). Based on high diversity, complex molecular structure(s), broad-spectrum activity including inhibition of viral transcription and translation, as well as considering their overall safety and non-cytotoxicity, phytochemicals may serve as promising anti-COVID-19 interventions (Lalani and Poh, 2020; Chojnacka et al., 2020). For example, several plant-derived flavonoids are potential inhibitors of SARS-CoV-2 with comparable or higher potency as that of antiviral drug, remdesivir (Tahir ul Qamar et al., 2020). Such natural antiviral agents that target highly conserved SARS-CoV-2 replication enzymes (i.e., Mpro or RdRp) provide two advantages: (i) potential for broad-spectrum antiviral activity, and (ii) reduced risk of mutation-mediated drug resistance (Naidu et al., 2021a; 2021b).
7.2.1. Phyto-nutrient inhibitors of SARS-CoV-2 main protease (Mpro)
Mpro also known as the chymotrypsin-like protease (3CLpro), is one of the well characterized antiviral targets among CoV pathogens (Grum-Tokars et al., 2008). Along with the papain-like protease (PLpro), Mpro is critical to process polyproteins that are translated from the viral RNA (Hilgenfeld, 2014). Therefore, Mpro is considered as one of the key antiviral targets to combat COVID-19 (Joshi et al., 2020). Antiviral agents targeting the Mpro enzyme could significantly reduce the risk of mutation-mediated drug resistance and display broad-spectrum antiviral activity (Goyal and Goyal, 2020). The phytochemical Mpro inhibitors could effectively target the large surface area of viral enzyme dimers (Hartini et al., 2021).
Medicinal herbs Isatis tinctoria, Torreya nucifera, Psoralea corylifolia, and Rheum palmatum could elicit potent anti-Mpro activity (Mandal et al., 2021). Baicalein, a natural compound commonly used in TCM, is a potent inhibitor of Mpro. In a RCT (n = 72), baicalein (100-2,800 mg) was well tolerated to treat acute, or chronic hepatitis (Li et al., 2014). The Mpro inhibitor NLC-001, a phytochemical supplement that is currently undergoing a human RCT in Israel, as a potential oral intervention against COVID-19 (Golodetz, 2021). Natural phytochemicals, pentagalloylglucose (PGG), and epigallocatechin-3-gallate (EGCG) demonstrate potent inhibition of Mpro (Chiou et al., 2021). Based on the pH milieu, Mpro could form dimers, tetramers, or even highly active octamers (Zhang et al., 2020b). Quercetin demonstrates a unique redox-based mechanism and inhibits Mpro by altering the thermostability of the enzyme (Abian et al., 2020). Quercetin is a promising scaffold to engineer new functional groups for the development of novel Mpro inhibitors.
7.2.2. Phyto-nutrient inhibitors of SARS-CoV-2 RNA-dependent RNA polymerase (RdRp)
RdRp also known as the ‘nsp12’, plays a central role in the replication and transcription cycles of SARS-CoV-2 (Gao et al., 2020). The RdRp enzyme polymerizes a high quantity of nucleotides to support an uninterrupted viral replication in the infected host (Naidu et al., 2022a). Due to its high evolutionary stability, RdRp has no counterpart in human cells; therefore, represents a unique antiviral target (Gordon et al., 2020; Vicenti et al., 2021). Several FDA-approved RdRp nucleotide analog inhibitors (NAIs) against RNA viral pathogens have been repurposed and evaluated for clinical management of COVID-19 (Chien et al., 2020). Furthermore, high through-put screenings and in-silico studies have also elucidated the anti-RdRp activity of several FDA-approved non-nucleotide analog inhibitor (NNAI) drugs (Tian et al., 2021). Unfortunately, the first open label, RCT with popular repurposed NAI drugs such as remdesivir, favipiravir, lopinavir-ritonavir, ribavirin, sofosbuvir, etc., showed poor efficacy against SARS-CoV-2 (Martinez, 2020). Considering the high morbidity/mortality rates of COVID-19 pandemic and lack of effective antiviral drugs, the repurposing of traditional antiviral phytonutrients and natural bioactives could be a promising strategy (Mahrosh and Mustafa, 2021a). A few phyto-nutrient RdRp inhibitors as potential intervention(s) for COVID-19 control are described below.
Suramin, a phytochemical from eastern white pine tree needles (Pinus strobus) is a potent NNAI against SARS-CoV-2 RdRp enzyme. A dosage of 100 µM suramin totally blocks the formation of RdRp-RNA complex compared to a 5 mM dosage of remdesivir; thus, the RdRp inhibition potency of suramin is at least 20-fold higher than remdesivir (Yin et al., 2020). Suramin also inhibits SARS-CoV-2 infection in cell cultures by blocking cellular entry of the virus (Salgado-Benvindo et al., 2020). Silibinin, a flavonolignan from the herb ‘milk thistle’ (Silybum marianum), directly inhibits the SARS-CoV-2 RdRp enzyme, and reduces the signal transducer and activator of transcription (STAT3)-induced lung and systemic inflammation in the infected host (Bosch-Barrera et al., 2020). The dual action of silibinin to inhibit viral replication machinery as well as host cytokine storm provides a strong rationale for clinical evaluation of this phytochemical for COVID-19 management.
Black tea polyphenols—theaflavin 3,3′-digallate (TF3), theaflavin 3-gallate (TF2a) and procyanidin B2 could effectively inhibit RdRp activity by blocking active site(s) in the enzymatic scaffold (Lung et al., 2020). Baicalein, a bioactive flavone found in the root of Scutellaria baicalensis, an ‘East Asian skullcap’ plant, could block viral replication in cell culture systems by inhibiting the RdRp activity (Song et al., 2021a; Zandi et al., 2021). Corilagin, is a gallotannin from plants Caesalpinia coriaria, Alchornea glandulosa and Punica granatum (pomegranate) bind directly to RdRp and inhibit the viral polymerase activity (Li et al., 2021b). Lycorine is a potent NNAI against SAR-CoV-2 RdRp enzyme with efficacy higher than the antiviral drug remdesivir (Jin et al., 2021). Propolis from (honeybee products) contains phenolics such as ellagic acid, hesperetin, and kaempferol that show high-affinity interactions with the SARS-CoV-2 RdRp and considered as effective COVID-19 inhibitors (Shaldam et al., 2021).
Apart from the NNAI spectrum these natural plant-based RdRp inhibitors also provide multifunctional dietary health benefits (Naidu et al., 2021a). Accordingly, these phytonutrients could provide synergistic or additive benefits with pharmaceutical drugs in immune modulation, anti-inflammation, and relieve oxidative stress in COVID-19 patients (Patil et al., 2021). Some predicted drugs and potential phytoceutical compounds that inhibit SARS-CoV-2 RdRp enzyme and modulate host physiological pathways are currently undergoing human clinical trials (Aronskyy et al., 2021).
7.3. Dietary/natural anti-inflammatories
Hyperinflammatory responses and inflammasome activation with SARS-CoV-2 infection are manifested as severe symptoms during COVID-19 (Pan et al., 2021). Once the ‘cytokine storm’ is triggered, just an anti-viral intervention alone is not sufficient to ameliorate the symptomatic progression of COVID-19. Additional anti-inflammatory treatments such as JAK inhibitors, IL-6 inhibitors, TNF-α inhibitors, colchicine, etc., may be necessary in the clinical management of the disease (Stebbing et al., 2020). The Nrf2/Keap1 signaling pathway mainly regulates anti-inflammatory gene expression, which could effectively contain SARS-CoV-2 infection (Cuadrado et al., 2020). Therefore, identification of new Nrf2-dependent anti-inflammatory phytochemicals has become a key investigative point in drug discovery (Ahmed et al., 2017).
The anti-inflammatory activity of several plant-based dietary nutrients could play a beneficial role in down-regulating the cytokine storm during COVID-19 (Liskova et al., 2021). Hesperidin, a glycosidic flavonoid from citrus species, could augment cellular antioxidant defenses via the ERK/Nrf2 signaling pathway (Parhiz et al., 2015). Hesperidin elicits anti-inflammatory effects via inhibition of the PI3K/AKT signaling pathway and suppress the secretion of proinflammatory cytokines such as INF-γ and IL-2, which may reduce the potential risks of cytokine storm in COVID-19 (Qi et al., 2019; Haggag et al., 2020). Phytochemicals Rutin and Punicalagin are also effective Nrf2 activators as well as potent SARS-CoV-2 Mpro inhibitors (Tian et al., 2016; Yan et al., 2016; Majumder and Mandal, 2020; Falade et al., 2021).
Quercetin is shown to inhibit inflammasome NLR family pyrin domain containing 3 (NLRP3) in macrophages (Saeedi-Boroujeni and Mahmoudian-Sani, 2021). Inflammasomes are cytosolic multi-protein complexes formed in response to cytosolic pathogen-associated molecular patterns (PAMPs) and damage-associated molecular patterns (DAMPs), which generate activated cytokines IL-1β and IL-18 (Zhao and Zhao, 2020). In a prospective RCT, quercetin supplementation (1-g daily/30 days) at an early stage of SARS-CoV-2 infection seem to reduce the frequency and duration of hospitalizations, the need for non-invasive O2 therapy, progression to ICU, and the CFR (Di Pierro et al., 2021). Several polyphenols (i.e., resveratrol, catechins), N-acetylcysteine (NAC), and palmitoyl-ethanolamide (PEA) could block inflammatory pathways (i.e., NLRP3 inflammasome-mediated IL-1β synthesis and pro-inflammatory cytokine release); as well as inhibit viral replication (by blocking the viral proteases (Mpro and RdRp) (Naidu et al., 2021a; 2021b). Myo-inositol, a naturally occurring polyol, widely found in foods, and actively synthesized in the body, seems to reduce IL-6 levels, and mitigate the inflammatory cascade (‘cytokine storm’), a prominent clinical manifestation of COVID-19 (Bizzarri et al., 2020).
7.4. Dietary/natural immune therapies
Optimal nutrition is essential for every cell in the body, including those of the immune system for an effective host defense (Childs et al., 2019). A ‘primed or activated’ immune system has a higher energy demands with increased basal metabolic rate (BMR) during the SARS-CoV-2 infection. Typically, the BMR increases by 10% during viral infection, accordingly the energy intake should be compensated during the illness (Kosmiski, 2011).
Both reactive and interactive states of host immune system plays a decisive role in the initiation, progression, severity, and fatality of a SARS-CoV-2 infection. Impaired Type I interferon (IFN-I) response leading to hyperinflammation (with ARDS and MODS) is a characteristic immune-pathobiology of COVID-19 (Mortaz et al., 2020), which is beyond the mechanistic explanation of viral tropism through the ACE2 and TMPRSS2 receptors. Interestingly, retinol insufficiency and impaired retinoid signaling are the intrinsic risk factors that may trigger immune dysregulation in COVID-19 (Sarohan et al., 2021). Immunotherapies with natural compounds that could help in inflammatory cytokine neutralization, immunomodulation, anti-inflammation and inflammation-associated systemic/organ damage could be potential adjuvant interventions in the management of acute COVID-19 or chronic PASC (Bonam et al., 2020). Several bioactive nutrients and phytochemicals have potential to boost and restore both innate and adaptive immune responses in COVID-19/PASC management. Therefore, immune-nutrition that could modulate inflammatory responses/cytokine release, viral antigen (wild epitopes or vaccines) recognition/processing and antibody production, could be an effective strategy for COVID-19 control.
Vitamin-A (retinol) levels in plasma drop to remarkably low levels during acute inflammation, which is evident in ARDS and mortality outcomes of COVID-19 patients (Tepasse et al., 2021). SARS-CoV-2 could suppress host type 1 interferons (IFN-I), the early innate immune response against viral infection. Therefore, replenishment of retinoid signaling pathways could be considered an effective strategy for management of COVID-19 as well as for treating chronic, degenerative, and inflammatory conditions in PASC. Retinoids (fat-soluble molecules related to vitamin A) are powerful immune-modulators capable of increasing IFN-I-mediated antiviral responses, which are currently investigated in several pre-clinical studies as adjuvants for antiviral drugs, vaccines and immune therapies against COVID-19 and PASC (Trasino, 2020).
Vitamin-A (Vit-A) plays a vital role in priming both innate and adaptive immune responses as well as in antibody production in COVID-19 patients (Midha et al., 2021). This fat-soluble vitamin regulates IL-2, TNF-α synthesis, activates macrophages (towards respiratory burst) and stimulates IFN-γ release to increase natural killer (NK) cell activity (Gombart et al., 2020). Mechanisms of action of Vit-A against SARS-CoV-2 include enrichment of immune reaction, containment of inflammatory reaction, and pathways related to ROS generation. Furthermore, seven core targets of Vit-A against COVID-19, including MAPK1, IL10, EGFR, ICAM1, MAPK14, CAT, and PRKCB have been reported (Li et al., 2020c).
Several natural compounds demonstrate antiviral activity via modulation of immune cell cascade (i.e., macrophages, dendritic cells), generation of cytotoxic (antiviral) factors such as nitric oxide (NO•), cytotoxic T lymphocytes from CD8+ T-lymphocytes, T-helper cells from CD4+ T-lymphocytes, activation of phagocytes, and proliferation of B-lymphocytes. (Jayawardena et al., 2020). Marine algae-derived bioactive compounds such as phlorotannins, alginates, luminaries, fucoidans, polyphenols, carotenoids, carrageenans and fatty acids are shown to enhance the human gut microbiota and revive health by enhancing epithelial barrier integrity, immune activity, and optimal metabolic function (Kumar et al., 2022). Milk proteins and peptides (i.e., lactoferrin (LF), lactoperoxidase (LPO), serum albumin, β-lactoglobulin, and α-lactalbumin) are well documented immune boosters (Naidu, 2000; Naidu et al., 2022b). Glycophosphopeptical (AM3) a natural glucan that modulates both innate and adaptive immunity, has also been suggested as an adjuvant for COVID-19 therapy (Fernández-Lázaro et al., 2021). Oral intake of glutamine (dosage: 0.3-0.5 g/kg/d), a conditionally essential amino acid, has been reported to modulate immune responses, which may alleviate clinical symptoms of cytokine storm during COVID-19 infection (Santos et al., 2020).
7.5. Dietary/natural antioxidants
Oxidative stress (OS) results from an imbalance between the formation of reactive oxygen species (ROS) and the innate antioxidant defense, which plays a crucial role in COVID-19 pathology (Cecchini and Cecchini, 2020; Suhail et al., 2020). The physiological levels of cellular free iron (Fe2+) must be tightly regulated to avoid ROS generation via the Fenton type-II reaction. In COVID-19 patients, the documented Fe-R-H alterations may cause iron overload, and trigger ferroptosis in multiple organs (Edeas et al., 2020). Fe-redox dysregulation (FeRD) with ensuing free radical (ROS) oxidative damage, is a common denominator in all symptomatic clinical phases of COVID-19 (Naidu et al., 2022a). Accordingly, the infected lungs COVID-19 patients show oxidized phospholipids, the degradation product of OS, which are clinical markers for acute lung injury (ALI) (Imai et al., 2008; Fratta Pasini et al., 2021). OS induced by NOX2 activation is linked with severe clinical outcome and thrombotic events in COVID-19 patients (Violi et al., 2020). SARS-CoV-2 infection seem to alter mitochondrial dynamics and trigger ROS production, with consequent OS, pro-inflammatory state, cytokine production, and cell death (de Las Heras et al., 2020). Such mitochondrial dysfunction may lead to different cellular responses such as adaptation to hypoxia, autophagy, inflammation, or ferroptosis. A decline in antioxidant defense, mainly the glutathione (GSH) with significant elevation in blood serum GSH reductase (from OS imbalance), has been observed in the ICU admitted COVID-19 patients (Cao et al., 2020). Furthermore, low levels of GSH show strong association with severe clinical outcomes and fatality rates in COVID-19 cases (Silvagno et al., 2020). Therefore, intervention strategies to reduce OS using antioxidants, NF-κB inhibitors, Nrf2 activators, and Fe-R-H regulators could be a promising approach for COVID-19 management.
7.5.1. Nutrient-based natural free radical scavengers/antioxidants
SARS-CoV-2 dysregulates the intracellular iron-redox homeostasis (Fe-R-H) state to manipulate host cell machinery for its advantage (Naidu et al., 2022a). Accordingly, Fe-R-H regulators are potential interventions to prevent both viral replication as well as virus-induced inflammation. Respiratory viruses, including SARS-CoV-2, significantly elevate ROS production and impair cellular defense systems; therefore, free radical scavengers could be considered as potential antiviral interventions (Checconi et al., 2020).
N-acetylcysteine (NAC) is a natural antioxidant derived from plants especially from the Allium species (Šalamon et al., 2019). As a precursor of reduced-GSH and direct ROS scavenger, NAC affects several cell-signaling pathways that regulate Fe-R-H, apoptosis, angiogenesis, cell growth and inflammatory response (Shi and Puyo, 2020). NAC could reduce intracellular hydrogen peroxide levels, prevent the OS-mediated activation of NF-κB and abrogate the upregulation of pro-inflammatory genes (Sadowska, 2012). NAC administration (oral, nebulization, or inhalation) has been suggested to interfere in OS-mediated endothelial damage and prevent micro-thrombosis in severe forms of the COVID-19 (Guglielmetti et al., 2020). The current experimental and clinical data suggests that NAC is a potential intervention against multiple therapeutic targets involved in the pathophysiology of SARS-CoV-2 infection (Fratta Pasini et al., 2021). However, these possible anti-COVID-19 mechanisms and properties of NAC need to be confirmed in human RCTs.
Glutathione (GSH) is the ‘master’ antioxidant in all tissues; the high GSH levels (reduced form) emphasizes its regulatory role in several biological processes including free radical scavenging, and resolving hyper-inflammation, the risk factors for multi-organ failure in COVID-19 (Polonikov, 2020). Intracellular GSH levels significantly decline during SARS-CoV-2 infection (Silvagno et al., 2020), and the GSH replenishment could effectively block viral replication through modulation of cellular redox state (Checconi et al., 2020). Therefore, GSH supplementation may provide an effective strategy to resolve inflammation and OS in COVID-19 and PASC patients (Guloyan et al., 2020). Oral administration of GSH (2,000 mg) is shown to relieve severe respiratory symptoms in COVID-19 patients (Horowitz et al., 2020). GSH intervention is a promising strategy; however, due to its poor bioavailability, the GSH dosage requirement for therapeutic efficacy is high. Accordingly, dietary supplementation with GSH precursors such as cysteine and glycine could be an alternate approach for effective COVID-19/PASC management (Silvagno et al., 2020).
Several dietary and plant-derived compounds are natural antioxidants capable of neutralizing or scavenging harmful free radicals, modulating several OS-mediated signaling pathways and protect cellular systems. Phytochemicals from Cat’s claw (Cadambine & Epiafzelechin-4β-8-epicatechin) and Ayurvedic medicinal herb ‘Tulsi’ (Isorientin 4′-O-glucoside) are active regulators of COX-mediated inflammatory pathways (Yuan et al., 2020). Quercetin is a naturally occurring plant flavanol from the flavonoid group of polyphenols, is a powerful scavenger of free radicals (i.e., ROS and RNS), hence classified as a redox nutraceutical known to reduce OS and inflammation (Xu et al., 2019).
Ergothioneine (ET) is a naturally occurring dietary amino acid and an intracellular antioxidant with metal ion affinity that accumulates to high levels in certain tissues in the body due to organic cation transporter novel-type 1 (OCTN1) (Gründemann et al., 2005; Halliwell et al., 2018). ET is shown to modulate inflammation, scavenge free radicals, counteract FeRD, prevent endothelial dysfunction, protect against ischemia, reperfusion injury, and ARDS. This broad-spectrum, multi-targeted therapeutic potential of ET could be beneficial in alleviating some major clinical manifestations of COVID-19 (Cheah and Halliwell, 2020). Furthermore, the ability of ET to cross the blood-brain barrier (BBB) and combat neuroinflammation, makes this natural antioxidant a promising candidate for intervention of both acute and chronic neuro-COVID symptoms (i.e., brain fog) during PASC (Koh et al., 2021). Human RCTs needs to be undertaken to establish the therapeutic and prophylactic potential of ET intervention against COVID-19/PASC management.
7.6. Dietary metabolic optimizers
COVID-19 patients, especially hospitalized individuals, show severe clinical manifestations, such as hypermetabolism and muscle catabolism (sarcopenia), due to acute systemic inflammation, with a reduced food intake leading to malnutrition. High oxidative stress is a characteristic of SARS-CoV-2 infection; accordingly, antioxidant supplementation may help reduce oxidative stress in high-risk populations (i.e., elderly and obese) and combat COVID-19. Adequate nutrition is vital for the proper cellular metabolism, and certain bioactive compounds such as essential amino acids, vitamins, and minerals, are critical for immune function and optimal metabolic function.
7.6.1. Micronutrients (Vitamins & Minerals)
Nutrition, especially micronutrients, plays an important role in reducing mortality and complications from COVID-19. The multi-functional role of micronutrients on host defense to contain SARS-CoV-2 infections would encourage several R&D strategies for prevention and control of acute COVID-19 as well as chronic PASC. Deficiency in micro-elements leads to progression of diseases and its replenishment to adequate levels could benefit patients with several protective factors such as immune boosters, help maintain redox homeostasis for better recovery from COVID-19. Nutrients such as vitamins A, C, D, zinc (Zn2+), and selenium (Se2+), through their synergistic adjuvant effects, could activate innate and adaptive immunity and counteract the progression of COVID-19 (Renata et al., 2022).
Vitamin-D (Vit-D), a widely studied prohormone associated with skeletal health, is a potent immunomodulator, with pleotropic effects on innate and adaptive immune cells (Kalia et al., 2021). Vit-D is shown reduce the synthesis of inflammatory cytokines (IL-2 and INF-γ) and elicit anti-inflammatory effects (Mansur et al., 2020). Vit-D receptors expressed on macrophages and other immune cells, regulate transcription of several genes, including those that produce antiviral peptides against SARS-CoV-2 (Kloc et al., 2021). Vit-D could affect ACE2 expression and regulate the host immune/vascular system through different mechanisms. It enhances the ACE2 to ACE ratio; thereby, increases angiotensin II hydrolysis, reduces subsequent inflammatory cytokine response and lung injury in COVID-19 (Rhodes et al., 2021). 1,25-dihydroxy-vitamin D (25OHD), the active form of Vit-D could protect endothelial membrane stability.
7.6.1.1. Vit-D deficiency (hypovitaminosis-D) and COVID-19
Vit-D deficiency is a high-risk factor for morbidity and mortality in COVID-19. Skin exposure to sunlight-UV stimulates Vit-D synthesis in the body. A significant correlation has been established between the geographical latitude of 88 countries and COVID-19 mortality rates, which corroborates with the observation that Vit-D deficiency contributes to COVID-19 pathobiology (Whittemore, 2020). A report from five W.H.O. regions (except the African continent), showed that low serum 25OHD levels are linked to higher risk of SARS-CoV-2 infection (Margarucci et al., 2021). Also, Vit-D deficiency is strongly associated with COVID-19 case fatality rates (CFR) in Europe and Asia. In a population analysis of 47 countries, the prevalence of Vit-D deficiency ranged from 6.9 to 81.8% in EU countries and 2.0 to 87.5% in Asian countries (Sooriyaarachchi et al., 2021).
Vit-D deficiency is linked to high risk of invasive mechanical ventilation and/or death (IMV/D) and increased CFR. In a clinical study (n = 185), a total serum 25OHD level of <12 ng/mL (<30 nM) has been considered as Vit-D deficiency (Radujkovic et al., 2020). A meta-analysis of about 2-million adult population revealed that Vit-D deficiency/insufficiency increases susceptibility to SARS-CoV-2 infection and severe COVID-19 outcomes (Dissanayake et al., 2022). The prevalence of Vit-D deficiency was 58.4% in COVID-19 patients with elevated levels of serum inflammatory markers (i.e., IL-6, ferritin, TNFα) (Jain et al., 2020). This observation translates into higher CFR in COVID-19 patients. COVID-19 patients with vitamin D-deficiency (<8 ng/mL) were reportedly four times more prone to severe fatality outcomes (Rodríguez et al., 2020b). Low 25OHD levels may predispose COVID-19 patients to an increased 28-day mortality risk (Vassiliou et al., 2020). Vit-D deficiency is more common among obese and diabetic individuals, major risk groups for COVID-19. Also, a relationship between Vit-D status and the severity of COVID-19 has been reported among pregnant women (Seven et al., 2021).
7.6.1.2. Vit-D supplementation for COVID-19
Vit-D status could serve as a prognostic marker for SARS-CoV-2 infections; accordingly, a Vit-D supplemental plan can be recommended to improve the clinical outcomes in COVID-19 patients (Ünsal et al., 2021). A large, population-based cohort study from Spain has reported that patients supplemented with cholecalciferol (n = 108,343) or calcifediol (n = 134,703) achieved serum 25OHD levels ≥ 30 ng/mL with better COVID-19 outcomes (Oristrell et al., 2022). A clinical study from Turkey reported that Vit-D supplementation (n = 163) could increase serum 25OHD levels (>30 ng/mL) in COVID-19 patients within 2-weeks, shortened the hospital stay and reduced the CFR by 2.14 times in COVID-19 patients (Gönen et al., 2021). An open-label, multicenter RCT has reported the effect of Vit-D supplementation on the prognosis of COVID-19 in high-risk older patients (n = 260). High-dose cholecalciferol (200,000 IU/d) was found effective, well-tolerated, and readily accessible for COVID-19 treatment (Annweiler et al., 2020).
Treatment of hospitalized COVID-19 patients (n = 106) with 25OHD could correct the Vit-D deficiency/insufficiency, improve the immune function by elevating lymphocyte percentage in the blood and decreasing the ‘neutrophil-to-lymphocyte’ ratio (Maghbooli et al., 2021). The lower neutrophil-to-lymphocyte ratio is associated with reduced ICU stay and CFR in COVID-19. In a placebo-controlled RCT (n = 40), vitamin D-deficient COVID-19 patients receiving cholecalciferol (60,000 IU/d) for 7 days, turned RT-PCR negative for SARS-CoV-2 with a significant decrease in serum fibrinogen levels (Rastogi et al., 2022). Also, oral Vit-D supplementation (5,000 IU/d) for 2-weeks could effectively reduce the recovery time for cough and ageusia (sensory loss of taste) in Vit-D-deficient patients with mild to moderate COVID-19 symptoms (Sabico et al., 2021).
Individuals with hypovitaminosis-D are at higher risk of SARS-CoV-2 infection; therefore, should consider consuming Vit-D to maintain the circulating 25OHD at optimal levels (75-125nmol/L) to combat COVID-19. Hypovitaminosis-D affects over 1 billion people worldwide with severe implications on cardiovascular and inflammatory disorders. Since many clinical studies have reported a strong correlation between hypovitaminosis D and COVID-19, management of COVID-19 and PASC through Vit-D supplementation is a promising intervention strategy (Verdoia and de Luca, 2021). However, well-designed RCT studies with robust field data is warranted to establish the efficacy of Vit-D supplementation in COVID-19 and PASC control.
Vitamin-C (Vit-C), also known as L-ascorbic acid, is a water-soluble nutrient with potent antioxidant, anti-inflammatory activities and plays a major role as co-factor/modulator in various pathways of the immune system. Vit-C helps in normal neutrophil function, free radical scavenging, regeneration of vitamin E, modulation of signaling pathways, activation of pro-inflammatory transcription factors, activation of the signaling cascade, regulation of inflammatory mediators, phagocytosis and increasing the neutrophil motility to the site of infection (Uddin et al., 2021). Vit-C is a promising therapeutic candidate, both in prevention and amelioration of COVID-19 infection, and in adjunctive therapy for critical care of PASC.
7.6.1.3. Intravenous Vit-C for COVID-19
A meta-analysis of 12 RCTs (n = 1,766) showed that Vit-C could reduce the length of ICU stay on average by 8%, shorten the duration of mechanical ventilation in ICU patients and significantly reduce the CFR in sepsis patients (Hemilä and de Man, 2021). In a RCT (n = 120), Vit-C (500-mg/day for 2 weeks) supplementation resulted in a higher mean survival duration compared to the control group (Majidi et al., 2021). In another RCT, COVID-19 patients receiving intravenous Vit-C (50 mg/kg/day) along with standard therapy, turned symptom-free faster with reduced hospital stay, than patients on standard therapy only. A high dose of intravenous Vit-C may block several critical inflammatory pathways of cytokine storm, help improve pulmonary function and reduce the risk of ARDS of COVID-19 (Liu et al., 2020b). In a retrospective cohort study (n = 76), high-dose intravenous Vit-C (6-g twice on day-1, and 6-g once for the following 4 days) reduced the risk of 28-day mortality and improved the oxygen support status in COVID-19 patients (Gao et al., 2021). In a critically ill COVID-19 patients, intravenous sodium ascorbate (60 g) could restore arterial pressure, improve renal function and increase arterial blood oxygen levels (May et al., 2021). COVID-19 patients (n = 55) administered with high-dose intravenous Vit-C (n = 55, 100 mg/kg/da) showed shorter duration of systemic inflammatory response syndrome (SIRS), lowered C-reactive protein levels and higher number of CD4+ T cells, compared to the control group. The levels of coagulation indicators, including activated partial thromboplastin time and D-dimer were also improved in the Vit-C treated group compared to the control (Zhao et al.,2021). Vit-C may prevent disease aggravation in the early stage of COVID-19, which may be related to improved inflammatory response, immune function, and coagulation function. Intravenous Vit-C administration may reduce the need for vasopressor support in hospitalized COVID-19 patients. The use of high dose of intravenous Vit-C for management of COVID-19 in China and the United States has shown promising results.
7.6.1.4. Vit-C status in COVID-19 pandemic
Supplementation with micronutrients, such as vitamins and minerals, has gained an increasing interest as part of the supportive management for COVID-19. Serum Vit-C levels and leukocytes are depleted during the acute stage of infection due to increased metabolic demand; therefore, high-dose Vit-C helps normalize such clinical condition (Abobaker et al., 2020). Patients with hypovitaminosis C or severe respiratory illnesses, such as COVID-19, may benefit from taking Vit-C, due to its well documented safety profile, simplicity of use, and potential health benefits. Oral Vit-C, when taken at onset of SARS-CoV-2 infection, may reduce the duration of symptoms including fever, chest pain, chills and bodily aches and pains (Schloss et al., 2020). Vit-C represents an appealing option for clinical management of the global COVID-19 pandemic due to its clinical efficacy, relative safety, and low cost. However, there is an absence of high quality, contemporary RCTs to better understand the value of both oral and IV vitamin C for COVID-19 management.
Mineral - Zinc (Zn2+) is the second most abundant trace element in the human body, an integral component of several enzymes and transcription factors in human physiology. Zn2+ is critical for maintaining the natural tissue barrier function (such as in the respiratory epithelium) and establish redox homeostasis to block cellular entry and detrimental progression viral pathogens, such as SARS-CoV-2 (Wessels et al., 2020). The anti-COVID spectrum of Zn2+ revolves around its ability to inhibit viral enzymes—RNA-dependent RNA polymerase (RdRp) and Mpro (Panchariya et al., 2021). Zn2+ also down-regulates ACE2, the zinc metallo-enzyme that serves as host cellular receptor for SARS-CoV-2 infection (Karim et al., 2021). The immune-regulatory activity of Zn2+ includes antiviral protection via up-regulation of interferon-α (IFN-α) synthesis, elicit anti-inflammatory activity by inhibiting NF-κB signaling and by modulating the regulatory T-cell functions that could limit the ‘cytokine storm’ in COVID–19 (Skalny et al., 2020). Additionally, Zn2+ could protect against hypoxic damage, a detrimental end-organ injury in ARDS (Perera et al., 2020). Therefore, Zn2+ is considered a promising prophylactic and therapeutic adjuvant for COVID-19 control, that helps reducing inflammation, improving muco-ciliary clearance, preventing ventilator-induced lung injury, and modulating antiviral immunity (Skalny et al., 2021).
7.6.1.5. Zn2+ deficiency in COVID-19
A strong correlation exists between low zinc levels and severe ARDS in critically ill COVID-19 patients (Gonçalves et al., 2021). Zn2+ deficiency has been attributed with the insurgence and progression of COVID-19 in the elderly population with severe clinical outcomes with increased risk of ARDS (Coni et al., 2021). Zn2+ deficiency alters ACE2 activity, which may lead to a hyper-inflammatory response, vasoconstriction and pro-thrombotic events (Gouda et al., 2021).Furthermore, several widely used anti-hypertensive drugs and statins could negatively affect Zn2+ status, impair antiviral immunity, increase susceptibility to SARS-CoV-2 infection (Mossink, 2020). Therefore, Zn2+ supplementation should be considered a prophylactic measure for such COVID-19 risk groups.
Free Zn2+ is an exchangeable and bioactive form of metallic ion in serum, a reliable biomarker for alterations in zinc-homeostasis and may serve as a prognostic tool to measure severity and course of COVID-19. Accordingly, SARS-CoV-2 infection is characterized by a marked decline in total serum Zn2+ levels and such deficiency is associated with increased risk of mortality in COVID-19 patients (Maares et al., 2022). In a hospital study, COVID-19 patients showed significantly lower Zn2+ levels (n = 47; median ∼75 μg/dL) compared to the healthy controls (n = 45; median ∼105 μg/dL). Such patients exhibit severe clinical manifestations including ARDS, extended hospital stay, and increased mortality (Jothimani et al., 2020). A cohort study from Belgium (n = 139) reported that an absolute majority of COVID-19 patients (96%) were deficient with Zn2+ (<80 µg/dL) (Verschelden et al., 2021). Such Zn2+ deficiency appears to be associated with impaired antiviral response, cytokine releasing syndrome (CRS), and ARDS, which are prominent during severe COVID-19.
7.6.1.6. Zn2+ supplementation for COVID-19
A recent meta-analysis (5 studies with 1,506 participants) suggested that Zn2+ supplementation is associated with a lower mortality rate in COVID-19 patients (Tabatabaeizadeh, 2022). In another RCT, patients (n = 164) administered with zinc sulfate as adjunctive therapy showed a low 30-day mortality and this additional treatment in critically ill COVID-19 patients seem to improve survival rates (Al Sulaiman et al., 2021). Cohort studies and RCTs are further warranted to establish robust evidence on the association between individual Zn2+ status and COVID-19 pathophysiology.
Mineral - Selenium (Se2+) is one of the major trace elements required for redox homeostasis related to immune-endocrine function and signaling transduction pathways (Gorini et al., 2021). Se2+ is incorporated into seleno-proteins with broad protective functions that may reduce the incidence and severity of COVID-19 (Zhang et al., 2020a). Se2+, more specifically the amino acid seleno-cysteine, is present in at least 25 human seleno-proteins involved in several vital biological functions, ranging from free radical scavenging to hormone biosynthesis (Barchielli et al., 2022). Accordingly, Se2+ is a cofactor for several antioxidant enzymes such as Glutathione peroxidases (GPx) that regulates inflammatory responses in the body (Kieliszek, 2021). In the seleno-enzyme GPx, both the functional complexes—Se2+ and GSH, correlate with clinical outcomes of SARS-CoV-2 infection. Any dietary deficiencies of Se2+ and/or GSH precursors would increase oxidative stress and inflammation that may trigger potential damage to multi-organ systems (Taylor and Radding, 2020). The Se-transporter seleno-protein P (SELENOP), the Se2+ status biomarker, is particularly low (<2.6 mg/L) in COVID-19 patients (Heller et al., 2021).
7.6.1.7. Se2+ deficiency and COVID-19
The deficiency of Se2+ in COVID-19 patients correlates with disease severity and mortality risk (Demircan et al., 2022). Serum Se2+ levels are significantly lower in COVID-19 patients (∼78 μg/L) compared to healthy controls (∼92 μg/L) (Younesian et al., 2022). Also, Se2+ and SELENOP levels are significantly low in acute patients compared to post-COVID-19 patients (Skesters et al., 2022). Notably, COVID-19 survivors tended to have higher Se2+ levels after 10-14 days compared to non-survivors (Notz et al., 2021). The Se2+ deficiency is also associated with higher viral susceptibility and more severe clinical outcomes, especially in the elderly COVID-19 patients.
In a German study, serum samples from COVID-19 patients (n = 33) when compared to the reference data from a European cross-sectional analysis (EPIC, n = 1,915), patients showed a pronounced deficit in total serum Se2+ (<45.7 µg/L) and SELENOP (<2.56 mg/L) levels (Moghaddam et al., 2020). Notably, the Se2+ status was significantly higher in COVID survivors compared to non-survivors. The Se2+ status analysis seems to provide a prognostic tool and an adjuvant strategy for Se2+ supplementation to treat severely diseased and Se-deficient COVID-19 patients.
In a China study (14,045 cases from 147 cities) the CFR for COVID-19 showed an increase from 1.2% in non-Se-deficient areas, to 1.3% in moderate-Se-deficient areas, and further to 3.2% in severe-Se-deficient areas. Based on Se2+ content in ‘top soil’, the CFR gradually increased from 0.8% in non-Se-deficient areas, to 1.7% in moderate-Se-deficient areas, and further to 1.9% in severe-Se-deficient areas (Zhang et al., 2021b). The incidence of COVID-19 was >10-fold lower in Se-enriched than in Se-deficient regions in China (Liu et al., 2021a). Persistent Se2+ deficiency may further lead to infertility in adult population (Kieliszek, 2021).
7.6.1.8. Se2+ supplementation for COVID-19
Se2+ and seleno-proteins modulate host redox homeostasis, oxidative stress, and inflammatory responses (Bermano et al., 2021). Therefore, Se2+ supplementation could be beneficial to restore host antioxidant capacity, reduce apoptosis and endothelial cell damage as well as platelet aggregation. In severe phase of COVID-19, the host antioxidant system(s) exhaust while neutralizing toxic free radicals in the body; which is linked to a decline in Se2+ and SELENOP levels (Skesters et al., 2022). Administration (i.v) is shown to provide cytoprotective benefits in resolving clinical manifestations such as SIRS, septic shock, sepsis, ARDS, ventilator associated pneumonia (VAP); therefore, suggested as an adjuvant option for COVID-19 (Oliveira et al., 2022). Intravenous administration of selenite (1-mg) could elevate both Se2+ as well as SELENOP levels to normal range, and also increase GPx3 activity over time (Notz et al., 2021).
The Se2+deficiency could alter host defense cascade and allow viral infection by inducing OS and enabling viral mutagenicity that may amplify the virulence trait(s) of the viral pathogen to inflict severe damage to the host (Martinez et al., 2021). Therefore, Se2+-supplementation could provide an adjuvant support for vaccines and antiviral drugs in reducing the viral load, mutation frequency, and enhance herd immunity in populations with Se-deficiency. In summary, Se2+ supplementation at an appropriate dose could be an effective adjuvant therapy for SARS-CoV-2 infections. However, large cohort studies are warranted to verify the benefits of Se2+ supplementation for COVID-19 and PASC control.
7.6.2. Macronutrients (Omega-3 Fatty Acids and Probiotics)
A multi-variate analysis of food consumption practices in 170 countries on recovery from COVID-19 showed that several diets had a positive effect on patient recovery in developed countries, especially diets with higher content of lipids (e.g., omega fatty acids and PUFA), proteins, antioxidants (e.g., polyphenols and vitamin-C) and micronutrients (e.g., selenium and zinc). However, in countries with extreme poverty (high Global Hunger Index Score), foods provided minimal effect on recovery from COVID-19 (Cobre et al., 2021). The Mediterranean diet, Atlantic diet, and the Dietary Guidelines for Americans, are designed to provide essential vitamins, minerals, and phenolic compounds that could activate enzymatic and non-enzymatic antioxidant responses (Trujillo-Mayol et al., 2021). These balanced diets recommend the consumption of fruit, vegetables, legumes, olive oil, and whole grains with low intakes of processed foods and red meat. Personalized/targeted nutritional therapy needs to be initiated started in the early phase of SARS-CoV-2 infection and should be monitored till patient recovery for better clinical outcomes. Such nutritional strategy could prevent complications, support recovery, and help COVID-19 patients to achieve the best possible physical, functional, and mental health status.
Omega-3 fatty acids, specifically eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), play a role in inflammatory processes and immunomodulation for both innate and acquired immune systems (Rogero et al., 2020). Both DHA and EPA could affect the clumping of ‘toll-like receptors’ (TLR) in the cell membrane, prevent redox signals that activate NF-κB that produce pro-inflammatory mediators and help ameliorate complications of COVID-19 (Hathaway et al., 2020). In addition to antioxidant and anti-inflammatory effects, omega-3 fatty acids could regulate platelet homeostasis and lower the risk of thrombosis, a potential intervention for severe COVID-19 and PASC (Djuricic and Calder, 2021). Omega-3 supplementation could also block the viral entry by altering composition of fats in the bilipid membrane of cells (Weill et al., 2020).
Oral supplementation of EPA (1.5 g/d) and DHA (1.0 g/d) is shown to lower plasma levels of IL-6 and IL-1ß (Tan et al., 2018), the key inflammatory mediators involved in SARS-CoV-2-induced cytokine storm. A pilot study with COVID-19 patients (n = 100) reported lower mortality in individuals with high plasma levels of DHA and EPA (Asher et al., 2021). Elevated levels of EPA/DHA could reduce the membrane-bound arachidonic acid (Allaire et al., 2017), a precursor for synthesis of pro-inflammatory mediators (i.e., prostaglandins and leukotrienes). The presence of EPA/DHA in inflammatory cells could also block the activation of NF-κB, the key pro-inflammatory transcription factor, and abrogate the intracellular inflammatory cascade (Denys et al., 2005). Finally, EPA/DHA interaction with cell membrane phospholipids may disrupt lipid rafts, disassemble surface receptors, and block incoming inflammatory signals (Shaikh et al., 2015). These multifunctional effects of DHA/EPA in tandem could prevent the onset of cytokine storm and reduce the progression of COVID-19. The anti-inflammatory and immunomodulatory effects of DHA/EPA could prevent the onset of cytokine storm and may reduce COVID-19 progression and disease severity (Szabó et al., 2020).
Probiotics are live microorganisms, when administered in adequate amounts, confers health benefits to the host (Naidu et al., 1999). Probiotics and/or their metabolites may target either directly the viral pathogen and/or modulate the host immune system. Gut microbial metabolites transported via the circulation to the lungs could inhibit viral replication or improve the immune response against viral pathogens (Gautier et al., 2021). Intestinal dysbiosis could trigger exacerbated inflammation in the lungs and in the gut, which may eventually lead to increased CFR in several respiratory infections (de Oliveira et al., 2021). Accordingly, oral administration of probiotics could be a remedial approach to reestablish healthy gut microflora and control COVID-19.
7.6.2.1. The ‘gut-lung axis’ and COVID-19
The bidirectional interaction between the respiratory mucosa and the gut microbiota known as the ‘gut-lung axis’, could affect either prophylactic or pathologic immune responses against SARS-CoV-2 infection. High expression of ACE-2 receptors in the GI tract, severe manifestation GI symptoms, and abnormal changes to gut microbiome have raised concerns about GI involvement as a risk factor or as a potential modulator in the pathogenesis of COVID-19. Bringing homeostasis in the gut-lung axis could help reduce the mortality and morbidity rates associated with SARS-CoV-2 infection (Baradaran Ghavami et al., 2021). Additionally, the gut microbiota composition affects (and gets affected by) the ACE2 expression and contributes to the regulation of inflammatory responses. Furthermore, probiotics are shown to enhance vaccine efficacy against viral respiratory infections. SAR-CoV-2-induced gut dysbiosis may influence viral transmission and disease progression. An understanding of this pathobiology may design effective interventions (i.e., probiotics, diet modulation), which could potentially help combat the GI consequences of COVID-19 and PASC (Venegas-Borsellino et al., 2021).
7.6.2.2. Probiotic supplementation for COVID-19
A balanced gut microbiota and ACE2 expression could provide optimal therapeutic approaches for COVID-19 management, as well as maintains a symbiotic relationship of the microbiota with the host defense (Ahmadi Badi et al., 2021). About 19 ongoing RCTs are evaluating the efficacy of probiotics in COVID-19 prophylaxis and treatment (Chen and Vitetta, 2021b). A multi-center RTC (n = 100) reported that a 14-day supplementation of probiotic + enzyme compIex could resolve post-COVID-19 fatigue and improve patients’ functional status and quality of life (Rathi et al., 2021). In retrospective study (n = 123) from Wuhan-China, about 75.6% of COVID-19 patients treated with probiotics survived to hospital discharge, significantly longer than patients without probiotic therapy. The probiotic treatment did not affect the IL-6 levels; however, could modulate the immune responses and lower the incidence of secondary infections in COVID-19 patients (Li et al., 2021a). Their multi-functional health benefits, proven safety, easy dosage, economical and ready availability, makes probiotics as potential dietary adjuvants in COVID-19 and PASC management.
Adequate hydration (30 mL/kg actual body weight) is critical for total recovery of COVID-19 patients; therefore, these patients should increase their daily fluid intake (2.5–3 L/day) by consuming water, milk, fruit juice, broth, sports drinks, coffee, and tea (Wotton et al. 2008; Barrea et al., 2022).
| 8. Conclusion and future perspective | ▴Top |
From a pathobiology standpoint of COVID-19 – the emergence of new VOCs, rapid spread of the viral pathogen, the severity of infections, weighing on one side; while the other side, the absence of a prophylactic ‘multi-valent’ vaccine(s) and/or a potent therapeutic drug(s) able to inhibit the SARS-CoV-2 infection, there is a dire necessity to discover and develop effective intervention strategies to combat this ongoing deadly pandemic.
8.1. COVID-19 vaccine – current status
The ‘emergency use authorization (EUA) of vaccines has been considered an effective strategy combat COVID-19; however, the emergence of several SARS-CoV-2 ‘variants of concern’ has compromised the efficacy of both vaccines and monoclonal antibodies. Notably, most EUA vaccines are designed based on the S protein, which has a very high mutation frequency. Therefore, any variation of a single amino acid could affect the viral replication, transmission, or immune control evasion, and render the vaccine ineffective. To date, a prophylactic ‘multivalent vaccine’ against multiple SARS-CoV-2 variants is not available (Naidu et al., 2022a).
8.2. Repurposed anti-viral drug – current status
The W.H.O. Expert Panel recommended mortality trials (405 hospitals in 30 countries) of four repurposed antiviral drugs: remdesivir, hydroxychloroquine, lopinavir, and IFNβ-1a in hospitalized COVID-19 patients. Unfortunately, these drug regimens had little or no effect on the overall CFR, initiation of ventilation, and duration of hospital stay of these patients (WHO Solidarity Trial Consortium, 2021). Several multi-center RCTs and meta-analyses also support this observation (Indari et al., 2021, Martinez et al., 2021, Basu et al., 2022).
8.3. COVID-19 an HMR-related FeRD Syndrome?
The SARS-CoV-2 infection triggers host metabolic reprogramming (HMR) that could alter mitochondrial function with significant disruption in cellular bioenergy synthesis affecting several vital metabolic pathways for viral advantage. Consequently, the ensuing tri-phasic virulence spectrum, host susceptibility pattern(s), and symptomatic outcomes of COVID-19 clinically manifest as iron (Fe)-redox disruptive hematological syndromes. Accordingly, nutritional strategies, using Fe-R-H regulators, ferroptosis inhibitors, anticoagulants, and iron chelators may help ameliorate FeRD syndrome in COVID-19 patients (Naidu et al., 2022a).
8.4. Nutritional interventions for COVID-19/PASC
Altered nutritional status and malnutrition are related to HMR, several metabolic disorders, systemic inflammation, and deterioration of the immune system, which cumulatively compromise the host defense against COVID-19 and hamper patient recovery from the disease. Furthermore, the aging process could profoundly affect several physiological functions in the body, that eventually decreases the body’s ability to cope with stressors. Malnutrition could delay healing and increase the duration of hospitalization.
The World Health Organization (WHO) has emphasized the benefit of adequate nutrition to enhance the immune system, especially at the time of COVID-19 infection. The European Society for Clinical Nutrition and Metabolism (ESPEN) gave concise guidelines for an effective nutritional management of COVID-19 aimed at the prevention and treatment of malnutrition in these patients (Barazzoni et al., 2020). The Food and Agriculture Organization (FAO) of the UN, and European Food Information Council (EFC), Center for Disease Control and Prevention (CDC), Dietary Association of Australia (DAA) and United National Children’s Fund (UNICEF) have proposed various health/dietary guidelines and advice pertaining to COVID-19 pandemic. Guidelines and advice mostly include the consumption of fruits, vegetables, fish oil, probiotics, whole grains, cereals, and nuts. These foods could substantially improve the immune system and thereby elicit overall protection from SARS-CoV-2 infection and lower the risk of COVID-19 related health issues as well as improve the recovery of patients (WHO, 2020; de Faria Coelho-Ravagnani et al., 2021). Intake of functional foods/nutraceuticals and dietary supplements rich in various micronutrients with antioxidant, anti-inflammatory, and immunomodulatory properties could lower the risk of SARS-CoV-2 infection and help rapid recovery from COVID-19 (Wong et al., 2021; Chiu et al., 2021).
Finally, the evaluation of nutritional status should be mandatory for all COVID-19 and PASC patients. Such evaluation should be conducted at every stage of the clinical manifestation and recovery of COVID-19. Also, most patients experience weight reduction during COVID-19 infection, due to increased inflammation, appetite loss linked to anosmia/ageusia. Therefore, it is critical to correct the imbalance between energy expenditure and dietary (calorie) intake of such patients. In conclusion, the monitoring of nutritional status is of prime importance in the clinical management of hospitalized COVID-19 patients and in the rehabilitation of PASC individuals.
A. Satyanarayan Naidu (Conceptualization, Writing-Original draft, Review/Editing, Visualization, & Preparation of Figures); Fereidoon Shahidi (Review); Chin-Kun Wang (Review); Kenji Sato (Review); Aman Wirakartakusumah (Review); Ogugua C. Aworh (Review); and, Roger A. Clemens (Conceptualization, Writing-Review/Editing).
| References | ▴Top |